LADA Niva Legend 3 дв. – Технические характеристики – Официальный сайт LADA
Колесная формула / ведущие колеса
Расположение двигателя
Тип кузова / количество дверей
Количество мест
Длина / ширина / высота, мм
База, мм
Колея передних / задних колес, мм
Дорожный просвет, мм
Объем багажного отделения в пассажирском / грузовом…
Код двигателя
Тип двигателя
Система питания
Количество, расположение цилиндров
Рабочий объем, куб. см
Максимальная мощность, кВт (л.с.) / об. мин.
Максимальный крутящий момент, Нм / об. мин.
мин.
Рекомендуемое топливо
Объем топливного бака, л
Максимальная скорость, км/ч
Время разгона 0-100 км/ч, с
Городской цикл, л/100 км
Загородный цикл, л/100 км
Смешанный цикл, л/100 км
Снаряженная масса, кг
Технически допустимая максимальная масса, кг
Максимальная масса прицепа без тормозной системы /…
Тип трансмиссии
Передаточное число главной передачи
Передняя
Задняя
Размерность
Габаритные размеры Нива ВАЗ 21213, 21214, 2131 lada 4×4
Примечание: см. так же «Контрольные точки и размеры кузова»
Габариты Lada 4×4 (3-х дверная)
Вид сверху
Размеры салона, багажника, проёмов, свесов и пр.
(размеры приведены для прежней и новой 2020 г. модификации)
L11 | h21 | B11 | H1 | L21 | h31 | B21 | L3 min/max1 | Нз1 | Вз, | h51 | V баг. | B41 | |
LADA 4×4 | 905/ | 990 | 1385 | 1145 | 640/ | 925 | 1365 | 585/1310 | 465 | 1000 | 870 | 216/ | 1045 |
LADA 4×4 | 985/ | 1020 | 1385 | 1165 | 570/ | 910 | 1365 | 585/1310 | 465 | 1000 | 870 | 216/ | 1045 |
H51 | H61 | M1, кг | М2, КГ | М (М1+М2), | S°1 град. | h21 | D1 | Wb1 | А1 | Е1 | Тп/Тз1 | |
LADA 4×4 | 745 | 1930 | 730 | 482 | 1212 | — / — | 325 | 3640 | 2200 | 1640 | 1680 | 1440/ |
LADA 4×4 | 745 | 1930 | 730 | 482 | 1212 | — / — | 325 | 3640 | 2200 | 1640 | 1680 | 1440/ |

5-и дверная
размеры салона и багажника
Габариты Lada 4×4 Urban (3 дв.)
5-и дверный Urban
Габаритные размеры (справочные) автомобилей ВАЗ-2129, -2130 и их модификаций
Габаритные размеры (справочные) автомобилей ВАЗ-2131, -21312 и их модификаций
Габариты Нива Ландоле (ВАЗ-212183, 212183-20)
Максимальный угол бокового наклона до опрокидывания (крена, 2121)
Lada Niva 4×4 5 дв.
 Urban 2021 обзор, комплектации и цены, характеристики
Urban 2021 обзор, комплектации и цены, характеристики Обзор LADA 4×4 Urban 5 дв.
LADA 4×4 Urban. Создана для приключений
Трудные дорожные условия. Теснота городских кварталов. Бескрайние поля и лесные тропинки… Это среда обитания LADA 4х4, она была, есть и будет. Именно поэтому LADA 4х4 входит в число самых легендарных автомобилей мира, вызывающих восхищение миллионов людей.
Отличное сочетание компактности, проходимости и комфортной цены – это делает LADA 4х4 необыкновенно популярной. Сохраняя выдающие внедорожные свойства и узнаваемый стиль, мы сделали автомобиль более комфортным, более функциональным и безопасным.
Добро пожаловать в мир LADA 4х4! Мир, в котором для вас открываются совершенно новые возможности!
Новая эргономичная панель приборов. Увеличенный вещевой ящик, а главное – более удобная система управления отоплением и вентиляцией с тремя вращающимися рукоятками и электроприводом заслонок отопителя.
Система экстренного оповещения обязательна для современных автомобилей.
Более комфортные передние кресла получили развитую боковую поддержку и новый механизм складывания.
Городской стиль LADA 4х4
Проходимость и высокая посадка LADA 4х4 востребованы и в крупных городах. Именно поэтому пользуется успехом пятидверный LADA 4×4 Urban – вместительный внедорожник с узнаваемым харизматичным дизайном.
Новый комфорт
Сегодня все модели семейства LADA 4х4 стали комфортней за счет заметных и скрытых изменений. В LADA 4х4 Urban, который ездит прежде всего по асфальту, они заметны особенно! Вместе с более комфортным интерьером пятидверный LADA 4х4 Urban получил противотуманные фары – два круглых светлячка отлично вписались в массив бампера.
Комплектации и цены
| Крепления для детских сидений ISOFIX | ||
| Дневные ходовые огни | ||
| Антиблокировочная система с электронным распределением тормозных сил (ABS, EBD) | ||
| Система вспомогательного торможения (BAS) | ||
| Новая комбинация приборов | ||
| Розетка 12V | ||
| Розетка 12V в багажном отделении | ||
| Дополнительная виброизоляция | ||
| Гидроусилитель рулевого управления | ||
| Легкая тонировка стекол | ||
| Электростеклоподъемники передних дверей | ||
| Подогрев передних сидений | ||
| Электропривод и обогрев наружных зеркал | ||
| Кондиционер | ||
| Аудиоподготовка | ||
| Молдинги боковых дверей | ||
| Накладки порогов пола | ||
| 16» легкосплавные диски Grizzly двухцветные с шинами Pirelli | ||
| Запасное стальное колесо временного использования 16» | ||
Характеристики
| Расположение двигателя | ||
| Длина / ширина / высота, мм | ||
| База, мм | ||
| Дорожный просвет, мм | ||
Объем багажного отделения в пассажирском / грузовом. .. .. | ||
| Колесная формула / ведущие… | ||
| Тип кузова / количество… | ||
| Количество мест | ||
| Колея передних / задних колес,… | ||
| Тип двигателя | ||
| Система питания | ||
| Рабочий объем, куб. см | ||
| Код двигателя | ||
| Количество, расположение… | ||
| Максимальная мощность, кВт (л.с.) / об…. | ||
| Максимальный крутящий момент, Нм / об…. | ||
| Рекомендуемое топливо | ||
| Максимальная скорость, км/ч | ||
| Время разгона 0-100 км/ч, с | ||
| Городской цикл, л/100 км | ||
| Загородный цикл, л/100 км | ||
| Смешанный цикл, л/100 км | ||
| Снаряженная масса, кг | ||
| Объем топливного бака, л | ||
| Технически допустимая максимальная масса,… | ||
| Максимальная масса прицепа без тормозной системы /… | ||
| Тип трансмиссии | ||
| Передаточное число главной… | ||
| Передняя | ||
| Задняя | ||
Фотографии
Экстерьер
Руководства по эксплуатации
Руководство по эксплуатации LADA 4х4 от 26.10.20
Руководство по эксплуатации LADA 4х4 от 07.07.20
Руководство по эксплуатации LADA 4х4 от 03.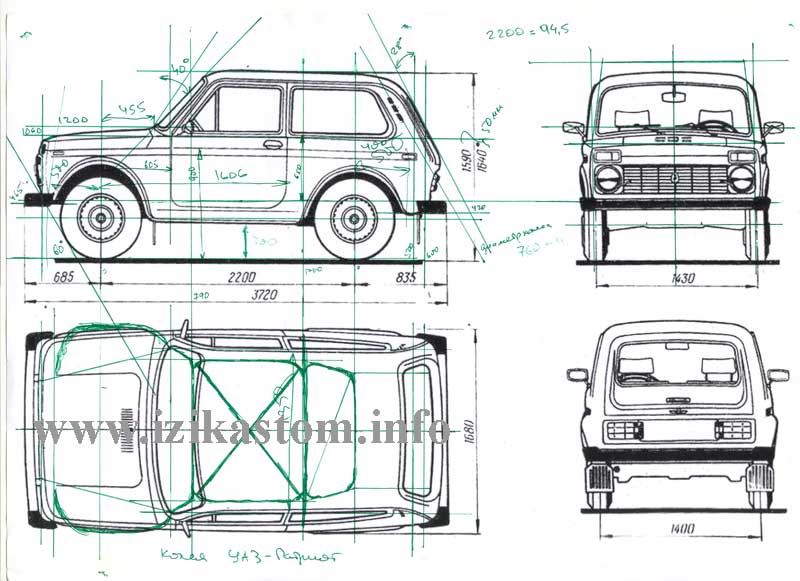 06.20
06.20
Руководство по эксплуатации LADA 4х4 от 24.03.20
Руководство по эксплуатации LADA 4х4 от 02.03.20
Руководство по эксплуатации LADA 4х4 от 20.01.20
Руководство по эксплуатации LADA 4х4 от 05.12.19
Руководство по эксплуатации LADA 4х4 от 04.09.19
Руководство по эксплуатации LADA 4х4 от 04.06.19
Руководство по эксплуатации LADA 4х4 от 07.09.18
Руководство по эксплуатации LADA 4х4 от 08.06.18
Руководство по эксплуатации LADA 4х4 от 30.05.18
Руководство по эксплуатации LADA 4х4 от 28.12.17
Руководство по эксплуатации LADA 4х4 от 08.06.17
Руководство по эксплуатации LADA 4х4 от 13.03.17
Руководство по эксплуатации LADA 4х4 от 31.01.17
Руководство по эксплуатации LADA 4х4 от 18.01.17
Руководство по эксплуатации LADA 4х4 от 23.12.16
Руководство по эксплуатации LADA 4х4 от 26.09.16
Руководство по эксплуатации LADA 4х4 от 26.05.16
Руководство по эксплуатации LADA 4х4 от 18.04.16
Руководство по эксплуатации LADA 4х4 от 07.04.16
Руководство по эксплуатации LADA 4х4 от 03.02.16
Руководство по эксплуатации LADA 4х4 от 25.01.16
Руководство по эксплуатации LADA 4х4 от 02.02.15
Руководство по эксплуатации LADA 4х4 от 12.12.14
Руководство по эксплуатации LADA 4х4 от 05.08.14
Руководство по эксплуатации LADA 4х4 от 12.09.11
Руководство по эксплуатации LADA 4х4 от 16.08.10
Руководство по эксплуатации LADA 4х4 от 25.06.08
Руководство по эксплуатации LADA 4х4 от 16.10.07
Руководство по эксплуатации LADA 4х4 от 10.10.07
Руководство по эксплуатации LADA 4х4 от 15.05.07
Руководство по эксплуатации LADA 4х4 от 22. 02.07
02.07
Тест-драйв Lada 4×4 Urban 5D: нужна ли сегодня в России «Нива»?
Вряд ли в далеких 70-х годах прошлого века два знаменитых советских инженера-конструктора — Владимир Сергеевич Соловьев и Петр Михайлович Прусов — могли себе представить, что их родное детище ВАЗ-2121 «Нива» когда-нибудь будет иметь название, в котором нет ни одного русского слова. А вот на то, что эта машина проживет сорок лет и останется на конвейере, тогда вполне можно было рассчитывать. Ну а что представляет собой эта легендарная модель сегодня: «Ниву» как ненужный пережиток прошлого или осовремененную классику Lada 4х4 Urban, способную дать бой сегодняшним недокроссоверам?
Конечно, никакие пластиковые бамперы, накладки на двери и новые трехмерные эмблемы не скроют от глаз кузов нашей родной «Нивы» образца 1977 года, а вернее — 1993-го. Именно в начале 90-х на конвейере опытно-промышленного производства АвтоВАЗа был дан старт растянутой на 500 мм пятидверной версии.
Собственно, латинское словцо Urban — не более чем название комплектации (доступна и для трехдверки), в которую добавили почти все возможные опции и слегка украсили декором. А ведь ходили слухи, что версия «Урбан», то есть «городская», лишится внедорожных атрибутов, оставшись в виде кроссовера — просто с постоянным полным приводом. Пронесло!
По всем габаритам (4140х1680х1640 мм) Lada 4х4 проигрывает тому же Renault Duster, однако оказывается длиннее родственного «соседа» по конвейеру Chevrolet Niva. По современной системе классификации все три автомобиля можно отнести к внедорожникам субкомпактного класса «B» без всякого +. Кстати, военный УАЗ «Хантер» длиной всего 4100 мм, правда, по ширине и высоте больше двух метров. УАЗ «Патриот» со своими почти пятью метрами на фоне всех перечисленных — громадина
Прежде чем попасть в салон, придется проникнуться ностальгией: ключики — что от почтового ящика — остались у «Нивы» с советских времен, в которых, как вы понимаете, никаких центральных замков не было. А потому, открыв водительскую дверь, пассажиров в салон придется пускать по очереди давно забытым подтягиванием вверх замочных «грибочков». Ну и, конечно, под руками должен быть баллончик силикона: в первые же заморозки водительская личинка замерзла, а отогревшись, перестала плотно закрываться. В общем, сигнализацию с функцией центрального замка на все двери можно сразу прибавлять к стоимости — еще и потому, что закрывать все четыре двери иногда попросту забываешь.
А потому, открыв водительскую дверь, пассажиров в салон придется пускать по очереди давно забытым подтягиванием вверх замочных «грибочков». Ну и, конечно, под руками должен быть баллончик силикона: в первые же заморозки водительская личинка замерзла, а отогревшись, перестала плотно закрываться. В общем, сигнализацию с функцией центрального замка на все двери можно сразу прибавлять к стоимости — еще и потому, что закрывать все четыре двери иногда попросту забываешь.
За исключением руля, щитка приборов и сидений — внутри 1977 год, вернее — опять 1993-й. Вместе с появлением пятидверной версии был произведен и первый (!) за 16 лет рестайлинг. Классический «шестерочный» салон получил модернизацию, которая сейчас на фото. Интерьер, конечно, вызывает снисходительную улыбку, но как его оценивать по современным меркам — сложный вопрос.
Начнем с того, что в Lada 4х4 Urban 5D — язык сломаешь! — то есть, черт побери, в «Ниве», просторно. Несмотря на откровенно скромные габариты, внутри ничто и никто не мешает, поскольку отделка интерьера — это, по сути, просто прикрытый пластмассой изнутри прямоугольный кузов.
Прикрыт он, по большому счету, удобно, поскольку все необходимое на виду. Однако до громкого слова «эргономика» все-таки далеко: кнопка включения ближнего света — на печке, вентилятора печки — рядом с клавишей заднего дворника, которая рядом с неработающей клавишей обогрева заднего стекла. Кнопки стеклоподъемников — рядом с ручником. И это еще не считая старых добрых тонюсеньких советских рычажков слева от руля, которыми регулярно включаешь дальний вместо поворотника.
Главное новшество «Нивы» наряду с ABS, гидроусилителем и подогревом сидений — конечно, кондиционер! А вот магнитолы нет — прибавляйте к стоимости вместе с динамиками.
К плотным сиденьям с обозначенным профилем скорее привыкаешь, как и к посадке. Вполне приятный в толщине и хвате руль, увы, не регулируется, само кресло — по двум направлениям и то специфически: салазки стоят под уклон назад, а сидушка, наоборот, вниз.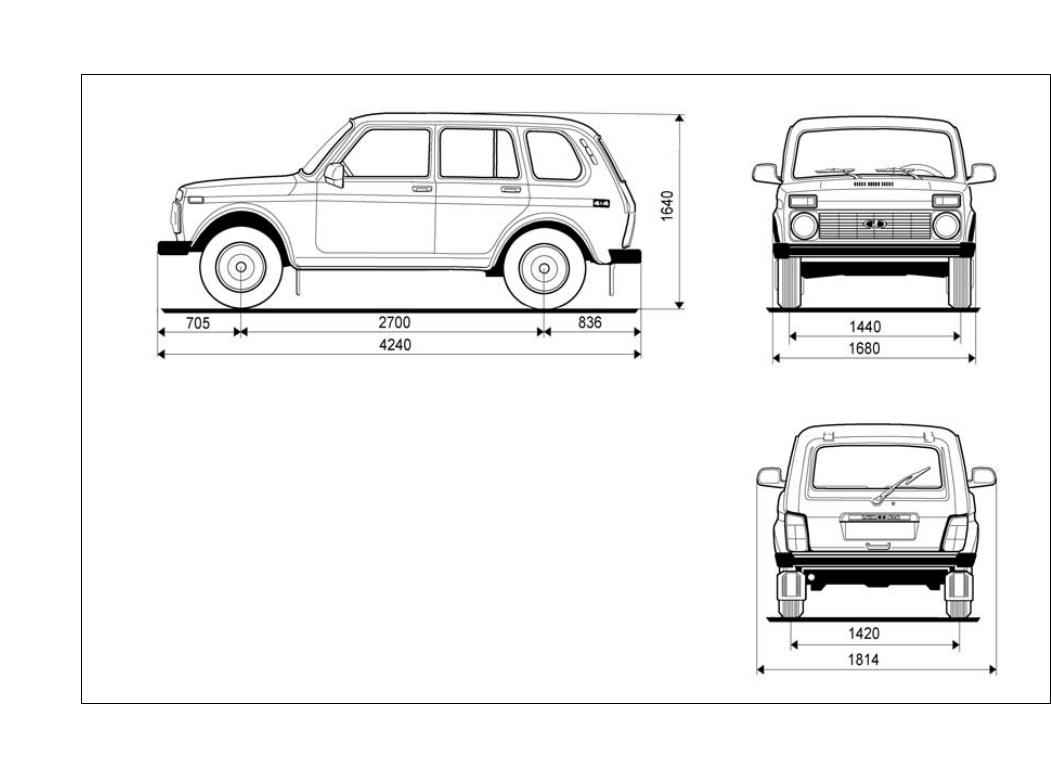 При этом настроиться придется не только по рулю и педалям, но и по рычагу КПП, до которого приходится тянуться. В общем, найти компромисс смогут люди среднего роста, да и то не все. А вот сзади, в общем-то, полный порядок: места достаточно даже для троих, благо пол почти ровный.
При этом настроиться придется не только по рулю и педалям, но и по рычагу КПП, до которого приходится тянуться. В общем, найти компромисс смогут люди среднего роста, да и то не все. А вот сзади, в общем-то, полный порядок: места достаточно даже для троих, благо пол почти ровный.
В «Ниве» осталось место и для приятных дополнительных благ. Есть карманы в дверях, подстаканники спереди, стакан-пепельница, своеобразная площадка для телефона рядом рычагами раздатки, полка под бардачком, карманы в спинках передних кресел, сеточки-держатели для замков ремней и даже крепления Isofix. Прогресс!
Не подводит и багажник: 420 литров полезного объема — это больше, чем в «Дастере». Причем задний диван складывается — правда, увы, только целиком. Наследство из прошлого — отдельный бачок омывателя для заднего стекла, из настоящего — розетка на 12 вольт
Под капотом родной, накрытый с рождения «запаской», вазовский мотор 1.7, «прокаченный» до норм ЕВРО-5 и 83 лошадиных сил. В паре с ним — все та же «семерочная» 5-ступенчатая механика, понижающая передача и возможность блокировки межосевого дифференциала системы постоянного полного привода — главной гордости машины.
Как все это передвигается в 2016 году? Начать надо с того, что различных технических доработок в «Ниве» было сделано множество, начиная с калибровки жесткости хода педалей или настроек работы мотора и заканчивая установкой того же усилителя руля. Так что если брать «Ниву» начала 90-х и сегодняшний Urban, даже без современных опций, шаг по ездовому комфорту будет огромный. Однако, как мы выяснили в ходе длительного теста, по тому, что можно еще улучшить, можно написать не одну кандидатскую.
Самое главное, в отличие от большинства современных автомобилей, «Нива» живая. Абсолютно все механизмы, которые могут издавать хоть какие-то звуки, — издают все. Гудит напропалую мотор, воет коробка передач (жаль, все время в одной тональности, а то можно было бы играть на ступенях мелодии), по сырости свистит соловьем какой-то из роликов, скрипят тормоза.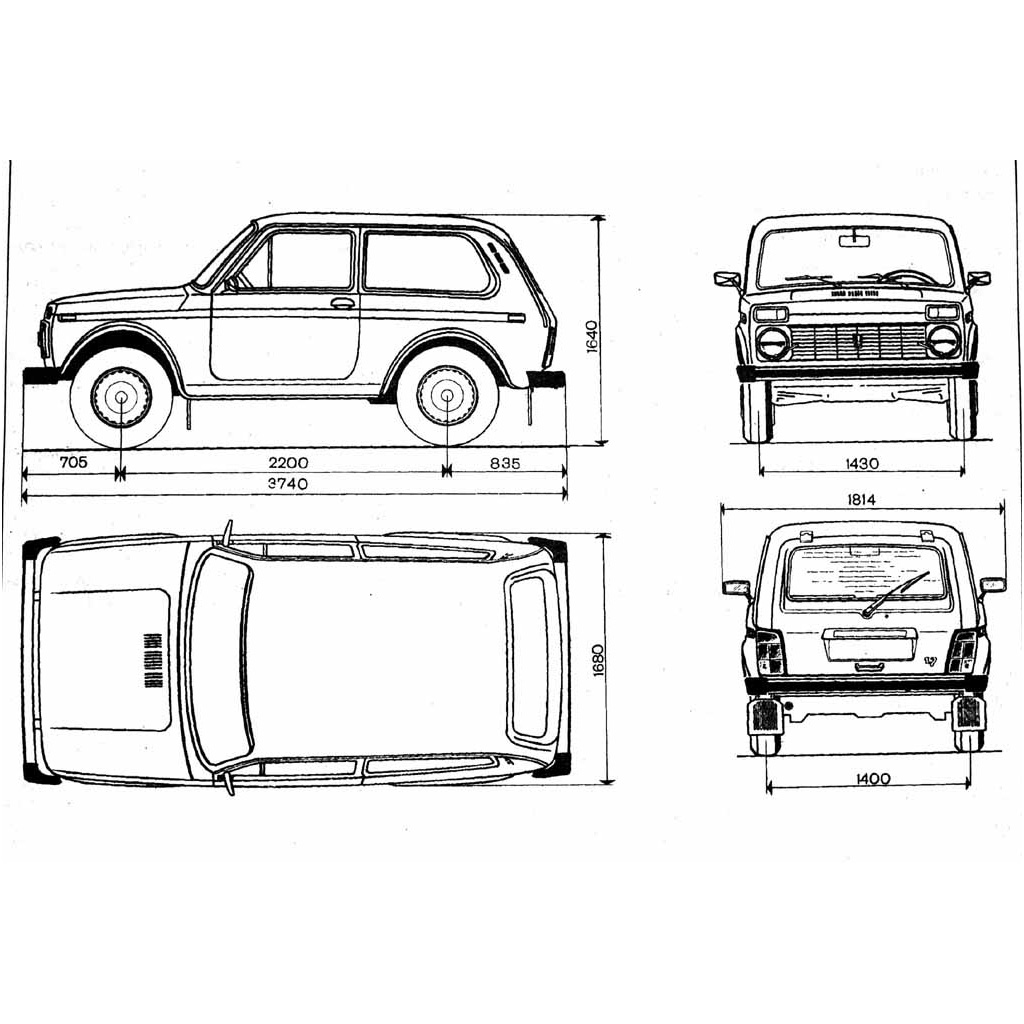 Даже стоя на светофоре, «Нива» чем-то тихо булькает в глубине мотора и периодически вздрагивает — так и хочется спросить: «Все ли хорошо, дорогая?». Все нормально! Потому что это русская баб… то есть машина. За три недели и почти 10 000 км общего пробега — ни одного вопроса, ни одной загоревшейся лампочки, ни одной серьезной поломки.
Даже стоя на светофоре, «Нива» чем-то тихо булькает в глубине мотора и периодически вздрагивает — так и хочется спросить: «Все ли хорошо, дорогая?». Все нормально! Потому что это русская баб… то есть машина. За три недели и почти 10 000 км общего пробега — ни одного вопроса, ни одной загоревшейся лампочки, ни одной серьезной поломки.
Огрехи сборки: почти сразу отвалились шайба регулятора фар, фиксатор сиденья и колесо регулировки его наклона, неплотно держащаяся эмблема на руле отковыривается одним движением. Мелочи, на скорость не влияет.
«Нива» не едет — по звукам и ощущениям, прет вперед. И, надо сказать, делает это вполне неплохо. Во-первых, несмотря на заявленные почти 20 секунд (!) до 100 км/ч, динамика до 50–60 км/ч, то есть городская, вполне конкурентоспособна: от потока машина особо не отстает, только успевай подтыкать передачи. Последние включаются четко, но размашисто: для пятой приходится отрываться от спинки кресла и заодно бить соседа по коленке. А вот выключать рычаг из пазов порой удается только легким ударом ладошки, как и втыкать заднюю — самую неинформативную ступень.
У длиннобазной «Нивы» вменяемые тормоза и великолепная курсовая устойчивость. Никаких рысканий и отклонений от курса при любой дороге и, главное, скорости — даже на предельных 140 км/ч управлять внедорожником без опаски можно одной рукой. Это, в том числе, заслуга универсальных всесезонно-внедорожных шин К-156-1 Кировского завода, которые на удивление, несмотря на свою узость, справляются даже с мокрой дорогой.
А вот с предельными нагрузками в поворотах не очень: слишком мягкие. В крутом вираже покрышки размерности 185/75/R16 подламываются почти до диска с угрозой повреждения и разбортирования. При этом сама машина неслабо кренится, хотя за рулем этого не чувствуется: склонности к заносу у «Нивы» нет, а снос выправляется сбросом скорости. Про подвеску и говорить нечего — тормозить перед лежачими полицейскими за время теста мы разучились. По энергоемкости амортизаторы не уступают дастеровским, а по плавности хода даже превосходят. Правда, сколько издевательств выдержат стойки — вопрос.
По энергоемкости амортизаторы не уступают дастеровским, а по плавности хода даже превосходят. Правда, сколько издевательств выдержат стойки — вопрос.
Съезд с асфальта для «Нивы» и ее водителя — праздник: оба могут резвиться в полную силу почти без ограничений. Собственно, внедорожные качества при компактных размерах и привлекательной цене — это то, за что «Ниву» любят и покупают.
Клиренс, хода подвесок, углы въезда/съезда и скрещивания осей, центральный «блок», «понижайка» и шины — полный арсенал почти для любых задач, которыми, правда, нужно уметь пользоваться. К примеру, с диагональным вывешиванием «Нива» борется только за счет блокировки. Любая грязь или ход в гору в натяг — только на понижающей передаче: из-за маломощного мотора приходится жечь сцепление. Ну и главное — длиннобазную версию есть большие шансы положить на «брюхо».
Каков итог?
Говорим как есть: если бы с подобным количеством доработок и опций машина вышла даже пускай в начале 90-х годов, это был бы компактный аналог тогдашнего какого-нибудь Mitsubishi Pajero — не меньше! Однако в современных условиях мегаполиса жить с Lada 4х4 Urban непросто. Если общие ездовые параметры автомобиля даже сегодня не вызывают особых проблем, то эксплуатационно-ездовые мелочи, которых набрался целый вагон, за три недели утомили.
«Нива», как и прежде, ежеминутно держит водителя в тонусе. Во-первых, это, конечно, безостановочный гул и свист всего вокруг, к которому со временем даже не привыкаешь, а просто смиряешься с ним. Во-вторых, мотор греется в пробках, отчего датчик температуры все время на контроле. Травит одно из колес, так что раз в два дня приходится его подкачивать. У печки всего два режима — либо ты поджариваешься, либо замерзаешь, и приноровиться к управлению системой микроклимата, чтобы получить что-то среднее, мы так и не смогли. Уже не работает обогрев заднего стекла. Из-за очень маленького угла поворота колес «Нива» не влезает в разворот даже на трехполосной дороге, не говоря уже о маневрах в рамках забитых дворов. Плюс все особенности эргономики сорокалетней давности.
Плюс все особенности эргономики сорокалетней давности.
Будь я реальным владельцем этой машины, на ближайшем вазовском сервисе меня бы уже, наверное, прокляли, учитывая, что ни одной поломки за 10 тыс. км пробега с машиной не произошло. Мы бы просто устраняли и усовершенствовали все то, что в новой «Ниве» попросту не доделано и недосмотрено, даже если это придется делать за собственный счет.
Покупать «Ниву» для города даже с еженедельными вылазками на дачу бессмысленно — только если дача находится в непролазных болотах.
Отсюда вывод. Для жителя современного мегаполиса «Нива» — это своеобразная игрушка. Явно не единственная машина в семье, которая выступает исключительно в виде тюнинг-проекта для серьезных внедорожных приключений — охоты, рыбалки или даже участия в трофи-рейдах. Учитывая, сколько подготовленных «Нив» я видел на различных внедорожных соревнованиях, спрос в этом сегменте хороший: дешевле из серьезных внедорожников попросту ничего нет.
Ну а второе применение, собственно, массовое: «Нива» — самый доступный, простой и, учитывая все доделки, относительно комфортный внедорожник для села, который ни на трассе, ни в городе не будет откровенным «утюгом». Даже отсутствие рядом фирменного сервиса вряд ли станет проблемой, поскольку родную «Ниву» наши мужики починят из копеечных запчастей и сами. Учитывая, что сегодня Lada 4х4 с сорокалетним бэкграундом входит в топ-20 самых продаваемых автомобилей России, отправлять легенду на покой действительно рано.
Шины, диски на Лада 4х4 (Lada 4×4)
Отечественный внедорожник Лада 4х4 славится тем, что может преодолеть любое бездорожье, особенно с правильно подобранными и установленными колесами. С разными размерами шин и колесных дисков этот автомобиль выглядит абсолютно по-разному. Со стандартными колесами небольшого диаметра он спокойно впишется в городской вид, а если у водителя есть желание или необходимость проложить дорогу там, где ее нет, для этой машины нет никаких границ.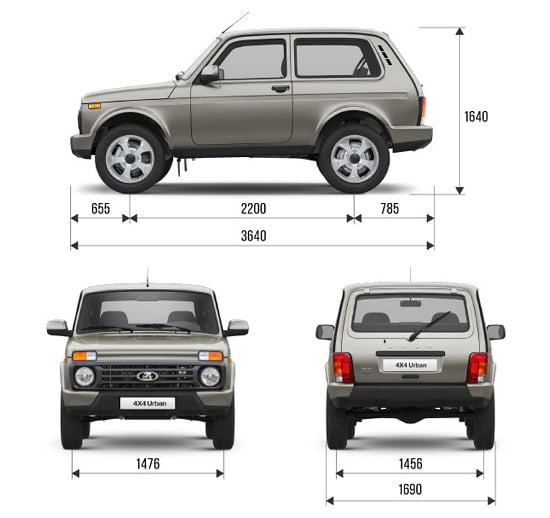
Главное, знать рекомендуемые размеры шин и дисков, определить свои ожидания от этого транспортного средства, узнать состояние дорог на местности, в которой он эксплуатируется.
Какие размеры шин и дисков рекомендуются в KOLOBOX для Lada 4×4?
Размер колесных дисков и шин зависит от года выпуска автомобиля и его модификации. Нива выпускается с 1984 года, является ветераном внедорожья. В таблице представлены размеры колес этого автомобиля в зависимости от перечисленных параметров.
Для внедорожника Лада Нива 4х4 возможны следующие размеры шин: диаметр 15 и 16 дюймов, ширина варьируется от 175 до 215 мм, а профиль (ширина к высоте) от 65 до 80 %. Но в зависимости от местности, в которой эксплуатируется этот автомобиль, водители устанавливают различные размеры.
Какая резина подходит для автомобиля Lada 4×4 в зимнее время года?
Многие автовладельцы автомобиля Лада 4х4 снимают штатные диски размера 16 дюймов и устанавливают 15-ти дюймовые для того, чтобы расширить выбор покрышек. В качестве 15- дюймовых дисков подходят диски от машины Нивы Шевроле. Монтаж возможен без доработок.
При выборе зимнего комплекта автопокрышек необходимо определиться с их типом: шипованные или фрикционные шины устанавливать? На этот вопрос многие владельцы Нивы отвечают просто: однозначно шипованные, т.к. автомобиль является внедорожным. Но решение должно быть индивидуально и зависит от погодных условий конкретного региона.
| Зимние автошины для авто Лада 4х4 | BFGoodrich G-Force Stud 102Q (XL)
Continental IceContact 2 SUV KD 102T (XL)(FR) Contyre Arctic Ice III 98T Cordiant Snow Cross (PW-2) 102T Michelin Agilis X-ice North |
|---|
Эти и многие другие модели можно установить на Лада 4х4 в зависимости от личных предпочтений.
Специалисты компании KOLOBOX ответят на все интересующие вопросы и подберут зимние покрышки для Нивы, которые будут отвечать всем необходимым требованиям.
Какие шины подходят для автомобиля Lada 4×4 в летнее время года?
Многие автовладельцы Лады 4х4 оснащают свой автомобиль различными вариантами шин, с которыми машина выглядит наиболее выигрышно и имеет высокий уровень проходимости.
С диаметром в 15 дюймов возможна установка покрышек размерностью 215/75 (ширина и профиль соответственно). Доработки при этом не требуются. Вылет диска должен быть ЕТ 35-58, а резина 27-28 дюймов. Если автомобиль будет перегружен или будет перемещаться по ухабистой местности, автопокрышки будут задевать колесные арки, что изнашивает покрышки.
Но есть и масса положительных сторон: потребление топлива снижается, увеличивается способность к преодолению любой местности, а в лифтинге не нужен.
Такая размерность представлена у множества брендов. Некоторые из них:
- Cordiant,
- Kumho,
- Michelin,
- Dunlop.
Какие диски рекомендуются для автомобиля Lada 4×4?
Для грамотного выбора колесных дисков для Лада 4х4 необходимо знать их параметры:
- Разболтовка вида: пять отверстий для крепления, находящиеся на окружности диска диаметром 139,7 мм.
- Вылет при размере шин 205/70 R15 — ЕТ 40, при 195/70 R15 — ЕТ 45, при формуле 175/80 R16 — ЕТ 48-58.
Какое давление рекомендуется для шин автомобиля Lada 4×4?
Завод-производитель указывает рекомендации, относительно давления в шинах Лада Нива 4х4, в зависимости от размеров колёс:
- Если шины этого автомобиля имеют формулу 175/80 (ширина и профиль соответственно) при диаметре 16 дюймов и 185/75 с тем же диаметром, давление в передних колесах должно быть 2,1 Атм, а в задних 1,9 Атм.
- Если покрышки имеют маркировку 195/70 с диаметром 15 дюймов и 205/70 с тем же диаметром, то давление в передних и задних колесах 1,9 Атм.
Каково влияние размера шин и дисков на характеристики автомобиля?
Рассмотрим влияние в таблице ниже:
Технические характеристики | АвтоСпецЦентр
Грузопассажирский пикап, выполненный на платформе легендарного внедорожника LADA 4×4. Имеет удлиненную двухдверную четырехместную кабину и грузовую закрытую платформу с откидным задним бортом.
Имеет удлиненную двухдверную четырехместную кабину и грузовую закрытую платформу с откидным задним бортом.
LADA 4×4 Пикап имеет более доступную по сравнению с аналогами стоимость
LADA 4×4 Пикап обладает высокой ремонтопригодностью
LADA 4×4 Пикап отличается высокой проходимостью
LADA 4×4 Пикап сочетает удобную кабину с грузовой платформой.
Габариты и масса
| тип кузова | пикап |
| габаритные размеры, длина/ширина/высота, мм | 4520×1680х1640 |
| вместимость, чел | 4 |
| масса снаряженного автомобиля, кг | 1420 |
| колея передних колес, мм | 1440 |
| колея задних колес, мм | 1420 |
| объем грузового отсека, куб. дм. | 620 |
| полная масса автомобиля, кг | 1810 |
Двигатель и трансмиссия Возможно исполнение с двигателем 1.8L
| тип | бензин, впрыск |
| рабочий объем, см³ | 1690 |
| мощность, кВт (л.с.) при об/мин | 61 при 5000 |
| крутящий момент, Н*м при об/мин | 129 при 4000 |
| нормы токсичности | Евро 4 |
| колесная формула | 4х4 |
| коробка передач, число передач | мех., 5-ступенчатая |
| раздаточная коробка, число передач | 2-ступенчатая |
| передаточное число главной пары | 3,9 |
| рулевое управление | травмобезопасное с телескопическим валом и гидроусилителем, рулевой механизм — глобоидальный червяк |
Комплектация Нива Пикап ВАЗ 2329 | Наличие |
Двигатель | 1,7 L |
Бензиновый, с распределенным впрыском топлива | + |
Безопасность |
|
Брусья безопасности дверей | + |
Иммобиллайзер | + |
Инерционные ремни безопасности | + |
Гидрокорректор света фар | + |
Экстерьер |
|
Увеличенные зеркала заднего вида с регулировкой из салона | + |
Пластиковая надстройка кузова | + |
Интерьер |
|
Обивка сидений – твид + искожа | + |
Регулировка уровня яркости подсветки органов управления | + |
Комфорт |
|
Гидроусилитель рулевого управления | + |
Очиститель-омыватель заднего стекла | + |
Багажники экспедиционные для ВАЗ 2121 Нива, Лада
Цена:
от: до:
Выберите категорию:
Все ТЮНИНГ УАЗ ТЮНИНГ НИВЫ И ШЕВРОЛЕ НИВЫ ТЮНИНГ ГАЗ СОБОЛЬ 4Х4 АККУМУЛЯТОРЫ И АКСЕССУАРЫ БАГАЖНИКИ ЭКСПЕДИЦИОННЫЕ, ЛЕСТНИЦЫ, РЕЙЛИНГИ» УАЗ ХАНТЕР, 469, 3151» УАЗ ПАТРИОТ» УАЗ БУХАНКА» ЛАДА / НИВА» ШЕВРОЛЕ НИВА» ГАЗЕЛЬ, СОБОЛЬ» LAND ROVER» TAGAZ TAGER» TOYOTA» NISSAN» MITSUBISHI» FORD / MAZDA» GREAT WALL HOVER» RENAULT DUSTER» SSANGYONG KYRON БАКИ ТОПЛИВНЫЕ БАМПЕРЫ СИЛОВЫЕ, ПОРОГИ И КОМПЛЕКТУЮЩИЕ СИЛОВОЕ ОБОРУДОВАНИЕ ARB БЛОКИРОВКИ БОДИЛИФТ (ЛИФТ КУЗОВА) БОКСЫ БАГАЖНЫЕ НА КРЫШУ ДИСКИ КОЛЕСНЫЕ И АКСЕССУАРЫ ДИСКОВЫЕ ТОРМОЗА ДОМКРАТЫ ЗАЩИТНЫЕ КЕЙСЫ ЗАЩИТА СИЛОВАЯ КАНИСТРЫ ЭКСПЕДИЦИОННЫЕ КОЛЕСНЫЕ МУФТЫ (ХАБЫ) КОМПРЕССОРЫ И ПНЕВМОСИСТЕМЫ КОНСОЛИ ПОТОЛОЧНЫЕ КУНГИ И АКСЕССУАРЫ ДЛЯ ПИКАПОВ ЛЕБЕДКИ И АКСЕССУАРЫ К НИМ ПОДВЕСКА, ЛИФТ-КОМПЛЕКТЫ ПРОСТАВКИ ДЛЯ УВЕЛИЧЕНИЯ КЛИРЕНСА РАСШИРИТЕЛИ КОЛЕСНЫХ АРОК РУЛЕВОЕ УПРАВЛЕНИЕ РУЛЕВЫЕ ДЕМПФЕРЫ И СТАБИЛИЗАТОРЫ СВЕТ ДОПОЛНИТЕЛЬНЫЙ СИНТЕТИЧЕСКИЙ ТРОС СНЕГОУБОРОЧНЫЕ ОТВАЛЫ СТРОПЫ, ТРОСЫ СЭНД-ТРАКИ ТОВАРЫ ДЛЯ АВТОТУРИЗМА УНИВЕРСАЛЬНЫЕ КРЕПЛЕНИЯ ФАРКОПЫ и КОМПЛЕКТУЮЩИЕ ЦЕПИ ПРОТИВОСКОЛЬЖЕНИЯ ШИНЫ ДЛЯ ВНЕДОРОЖНИКОВ ШНОРКЕЛИ ШТУРМАНСКОЕ ОБОРУДОВАНИЕ ЭЛЕКТРООБОРУДОВАНИЕ ПОДАРОЧНЫЕ СЕРТИФИКАТЫ
Производитель:
Все4REVO4WD SYSTEMS4x4sportAfcarfiberAfcarfiber (Турция)ALPHA (Тайланд)Amada XtremeARBArgus AnalyzersAVMBerkutBFGoodrichBiltemaBontyreBRCBUSHWACKER (США)CARRYBOYCHAMPIONChinaCM WinchCOME UPComforserCOOPERCORTLANDCSS (Турция)CSTDAKDarrDekaDelphiDelta(Германия)DuracellDynamica RopesEatonEGR (Австралия)End of The Road, Inc. EXIDEExpertDetalEXTANGFalconFARM JACKFatonFederalFlex LineGarminGOODYEARGT RadialHELLAHF DifferentialHi-liftIkon (Канада)INTERCOIRONMANJoker4x4K&KKAYMARKDT, РоссияKeko (Бразилия)KEMEIKing CobraKramcoKumhoKumho (Южная Корея)LAPTERLF WorksLightforceLinextras (Португалия)LOKKALongtengLS WheelsLUKMag LiteMarshalMaster-WinchMATECMaxlinerMAXTRAXMAXXISMechanix WearMetecMICKEY THOMPSONNankangNokian TyresOff-Road-WeelsOJ, РоссияOld Man EmuOPTIMAOptima BatteryORCIARIOTSOUTBACKPDWPirelliPitbul TiresPOWERFULProCompPROFORM (НОВАЯ ЗЕЛАНДИЯ)ProlightPT Group, ТольяттиQuattroRacing WheelsRALEX-TUNINGRanchoredBTRReplicaRivalROAD RANGER (Германия)Roll-N-Lock (США)RotopaxRuggedliner(США)RUNVARUSSACHSSAESAFARISAMMITR (Тайланд)SilverstoneSIMEXSJS (Тайланд)Sky-ComSM PowerSmittybiltStarcoSTARLEDSteelStaffSUPERSTONESuperwinchT-MAXTCC-tuningTecMateTELAWEITerratripTingerTopUp (Тайланд)TOPXENTorbikTough DogToyo (Япония)TplusTRIFFID TRUCKSTroffixTruXedoUFLEXUnielVAL-RACING, РоссияVFM-BosalVIAIRvnedorozhnik73.ruVoyagerWARNWelis LamontWincarX-Treme OutbackYokohamaYuBei Koyo Steering Systems Co.ZF, ГерманияАВС-ДизайнАВС-ДизайнАвтогур 73АвтоспасАвтоцепьАДСБАКОРБарнаул,РоссияБЗАКБлоккаБорисовВАКСОЙЛг. Курган, РоссияДАК, РоссияИЖ-ТЕХНО, РоссияИнсав, ТольяттиК&ККЗАТЭКиККРАМЗЛидерПлюсНИРФИ, РоссияНОМАКОННПК САМоВАРПолиуретан, РоссияПрофит+РИФРоссияРусская АртельСИБКОНТАКТСимбАТ, РоссияСОЮЗ-96СПРУТ, РоссияСтерлитамакСТОКРАТТайваньТайландТехно СфераТехноХимУльяновскШток Авто
EXIDEExpertDetalEXTANGFalconFARM JACKFatonFederalFlex LineGarminGOODYEARGT RadialHELLAHF DifferentialHi-liftIkon (Канада)INTERCOIRONMANJoker4x4K&KKAYMARKDT, РоссияKeko (Бразилия)KEMEIKing CobraKramcoKumhoKumho (Южная Корея)LAPTERLF WorksLightforceLinextras (Португалия)LOKKALongtengLS WheelsLUKMag LiteMarshalMaster-WinchMATECMaxlinerMAXTRAXMAXXISMechanix WearMetecMICKEY THOMPSONNankangNokian TyresOff-Road-WeelsOJ, РоссияOld Man EmuOPTIMAOptima BatteryORCIARIOTSOUTBACKPDWPirelliPitbul TiresPOWERFULProCompPROFORM (НОВАЯ ЗЕЛАНДИЯ)ProlightPT Group, ТольяттиQuattroRacing WheelsRALEX-TUNINGRanchoredBTRReplicaRivalROAD RANGER (Германия)Roll-N-Lock (США)RotopaxRuggedliner(США)RUNVARUSSACHSSAESAFARISAMMITR (Тайланд)SilverstoneSIMEXSJS (Тайланд)Sky-ComSM PowerSmittybiltStarcoSTARLEDSteelStaffSUPERSTONESuperwinchT-MAXTCC-tuningTecMateTELAWEITerratripTingerTopUp (Тайланд)TOPXENTorbikTough DogToyo (Япония)TplusTRIFFID TRUCKSTroffixTruXedoUFLEXUnielVAL-RACING, РоссияVFM-BosalVIAIRvnedorozhnik73.ruVoyagerWARNWelis LamontWincarX-Treme OutbackYokohamaYuBei Koyo Steering Systems Co.ZF, ГерманияАВС-ДизайнАВС-ДизайнАвтогур 73АвтоспасАвтоцепьАДСБАКОРБарнаул,РоссияБЗАКБлоккаБорисовВАКСОЙЛг. Курган, РоссияДАК, РоссияИЖ-ТЕХНО, РоссияИнсав, ТольяттиК&ККЗАТЭКиККРАМЗЛидерПлюсНИРФИ, РоссияНОМАКОННПК САМоВАРПолиуретан, РоссияПрофит+РИФРоссияРусская АртельСИБКОНТАКТСимбАТ, РоссияСОЮЗ-96СПРУТ, РоссияСтерлитамакСТОКРАТТайваньТайландТехно СфераТехноХимУльяновскШток Авто
Новинка:
Всенетда
Спецпредложение:
Всенетда
Результатов на странице:
5203550658095
2×4 Гитара: 10 ступеней (с изображениями)
Как и на любой другой гитаре, вам необходимо выполнить стандартную процедуру настройки. Отрегулируйте анкерный стержень для разгрузки шеи, установите высоту струны и настройку интонации, и все будет в порядке.
Удачи!
Edit: Некоторые люди спрашивают о стоимости. Я не хочу загромождать первую страницу кучей заметок, поэтому помещу ее сюда.
Я не хочу загромождать первую страницу кучей заметок, поэтому помещу ее сюда.
Я уже построил пару гитар раньше, так что у меня уже были некоторые детали и расходные материалы, так что это не точный подсчет того, сколько это мне стоило, а примерно столько, сколько это будет стоить тому, у кого есть большая часть инструментов и обычных товаров им доступны и нужны как раз и гитарные части, и дерево.Я отмечу некоторые альтернативы, где вы также можете сэкономить немного.
Пикапы 55 $
Купил активные пикапы GFS. Минимальным / более дешевым решением будет сборка с одинарным бридж-хамбакером, который стоит 20-30 долларов
Бридж 26 долларов
Базовый мост с креплением для хамбакера стоит около 17 долларов
Тюнеры 30 долларов
Pickguard 15 долларов
Горшки 12
Джек $ 11
Я пошел с хорошим запорным домкратом коммутатора. Вы можете приобрести стандартный разъем, который устанавливается на панель управления, примерно за 3 доллара США,
Ручки 11 долларов США
Это для набора стандартных ручек.Я потратил 13 долларов на несколько штук, покрытых морским ушком, сломал раковину, заменил ее углеродным волокном и накрыл эпоксидным куполом.
Caps 2 $
Switch 7 $
Я купил хороший трехпозиционный переключатель от digikey, потому что они нравятся больше, чем стандартные гитарные переключатели, но цена аналогична.
Ferrules 8 $
вы можете пропустить их, если у вас есть мост с отверстиями для струн на конце.
Контрольная пластина 14 $
Это довольно дорого для куска металла.Я просто ленился, когда заказывал. Сделайте один из металлолома, если хотите сэкономить несколько долларов.
Fretwire $ 10
4 фута должно быть достаточно, чтобы сделать любую разумную гитару или бас. Вероятно, этого достаточно для пары стандартных гитар, но неплохо иметь запасные на случай, если вы напортачите.
Лобовая пила 30 долларов
Обычная проволока для ладов подходит для паза 0,023 дюйма. Подойдет и немного шире, если вы собираетесь приклеивать лады. вы делаете гриф с плоским радиусом, и я хочу получить в будущем некоторые из дорогих алмазных накладки, но они делают свою работу.вы также можете обойтись без него, если вы действительно хорошо поработали над построением грифа и грифа и / или не возражаете против немного более высокого действия.
Подойдет и немного шире, если вы собираетесь приклеивать лады. вы делаете гриф с плоским радиусом, и я хочу получить в будущем некоторые из дорогих алмазных накладки, но они делают свою работу.вы также можете обойтись без него, если вы действительно хорошо поработали над построением грифа и грифа и / или не возражаете против немного более высокого действия.
Анкерный стержень 17 $
Играйте с регулируемым стержнем двойного действия (например, хот-род Stewmac и т. Д.). Избегайте компрессионных стержней и нерегулируемых стержней. Единственная причина, по которой их больше используют, — это «традиция»
Пуговицы на ремешке 5 долларов
Мне очень нравятся страплоки Dunlop, которые стоят около 20 долларов, но обычные тоже работают.
Дерево $ 10 + 12
Стоимость древесины довольно незначительна, если вы строите из пиломатериалов хвойных пород, как я.Он достаточно жесткий и звучит великолепно, поэтому не верьте никому, кто говорит, что он будет приглушенным или с отсутствием сустейна. Если вам нужны доказательства, поищите на YouTube сосновые тележки. В автомобилях 2х4 используется много разных сортов дерева. Те, что находятся в более тяжелом конце спектра, имеют жесткость, сопоставимую с кленом, и лишь немного хуже, чем красное дерево. (два наиболее распространенных вида дерева для шеи) В принципе, на нем будет немного легче вмятины, если вы ударите им что-нибудь, но в остальном это нормально.Убедитесь, что вы получили пиломатериалы, высушенные в печи. Если он станет зеленым, он покоробится, как крендель. Мне сказали, что даже KD все еще может деформироваться, но я построил эту гитару незадолго до Рождества 2013, и она все еще прямая (Может, удача, может быть, моя техника сборки?)
Итого: 315 долларов
Я не считал доставку , но в зависимости от того, из скольких мест вы заказываете, вы, вероятно, сможете оценить, сколько это будет вам стоить.
Новое поколение датчиков FRET для надежного измерения кинетики активации Gαi1, Gαi2 и Gαi3 в отдельных клетках
Abstract
Рецепторы, сопряженные с G-белком (GPCR), могут активировать гетеротримерный комплекс G-белка с субсекундной кинетикой. Генетически закодированные биосенсоры на основе резонансной передачи энергии Ферстера (FRET) идеально подходят для изучения таких быстрых сигнальных событий в отдельных живых клетках. Здесь мы сообщаем о создании и характеристике трех биосенсоров FRET для измерения активации Gα i1 , Gα i2 и Gα i3 . Чтобы обеспечить количественную долгосрочную визуализацию биосенсоров FRET с высоким динамическим диапазоном, необходимы флуоресцентные белки с улучшенными фотофизическими свойствами.Поэтому мы используем самый яркий и наиболее фотостабильный вариант CFP, mTurquoise2, в качестве донора, слитого с субъединицей Gα и , и cp173Venus, слитый с субъединицей Gγ 2 , в качестве акцептора. Конструкции биосенсоров Gα i FRET экспрессируются вместе с Gβ 1 из одной плазмиды, обеспечивая предпочтительные относительные уровни экспрессии с уменьшенными вариациями в клетках млекопитающих. Сенсоры FRET Gα и показали устойчивый ответ на активацию эндогенных или сверхэкспрессированных альфа-2А-адренорецепторов, которая ингибировалась токсином коклюша.Более того, мы наблюдали активацию датчика FRET Gα i в отдельных клетках при стимуляции нескольких GPCR, включая рецептор LPA 2 , M 3 и BK 2 . Кроме того, мы показываем, что датчики хорошо подходят для извлечения кинетических параметров из быстрых измерений в миллисекундном диапазоне времени. Это новое поколение биосенсоров FRET для активации Gα i1 , Gα i2 и Gα i3 будет полезно для измерений на живых клетках, которые проверяют активацию Gα i .
Генетически закодированные биосенсоры на основе резонансной передачи энергии Ферстера (FRET) идеально подходят для изучения таких быстрых сигнальных событий в отдельных живых клетках. Здесь мы сообщаем о создании и характеристике трех биосенсоров FRET для измерения активации Gα i1 , Gα i2 и Gα i3 . Чтобы обеспечить количественную долгосрочную визуализацию биосенсоров FRET с высоким динамическим диапазоном, необходимы флуоресцентные белки с улучшенными фотофизическими свойствами.Поэтому мы используем самый яркий и наиболее фотостабильный вариант CFP, mTurquoise2, в качестве донора, слитого с субъединицей Gα и , и cp173Venus, слитый с субъединицей Gγ 2 , в качестве акцептора. Конструкции биосенсоров Gα i FRET экспрессируются вместе с Gβ 1 из одной плазмиды, обеспечивая предпочтительные относительные уровни экспрессии с уменьшенными вариациями в клетках млекопитающих. Сенсоры FRET Gα и показали устойчивый ответ на активацию эндогенных или сверхэкспрессированных альфа-2А-адренорецепторов, которая ингибировалась токсином коклюша.Более того, мы наблюдали активацию датчика FRET Gα i в отдельных клетках при стимуляции нескольких GPCR, включая рецептор LPA 2 , M 3 и BK 2 . Кроме того, мы показываем, что датчики хорошо подходят для извлечения кинетических параметров из быстрых измерений в миллисекундном диапазоне времени. Это новое поколение биосенсоров FRET для активации Gα i1 , Gα i2 и Gα i3 будет полезно для измерений на живых клетках, которые проверяют активацию Gα i .
Образец цитирования: van Unen J, Stumpf AD, Schmid B, Reinhard NR, Hordijk PL, Hoffmann C, et al. (2016) Новое поколение датчиков FRET для надежного измерения кинетики активации Gα i1 , Gα i2 и Gα i3 в отдельных клетках. PLoS ONE 11 (1):
e0146789. https://doi.org/10.1371/journal.pone.0146789
https://doi.org/10.1371/journal.pone.0146789
Редактор: Микель Гарсиа-Маркос, Медицинский факультет Бостонского университета, США
Поступила: 25 сентября 2015 г .; Одобрена: 22 декабря 2015 г .; Опубликовано: 22 января 2016 г.
Авторские права: © 2016 van Unen et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: У авторов нет поддержки или финансирования, чтобы сообщить.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Подкласс Gα i гетеротримерных G-белков состоит из 3 членов у человека, Gα i1,2,3 , кодируемых генами GNAI1, GNAI2, GNAI3 [1] и активируемых широким диапазоном Рецепторы, сопряженные с G-белком. G-белки семейства Gα i участвуют в многочисленных патологиях, от участия в ожирении и диабете [2], функций иммунной системы [3] до их критических ролей на нескольких этапах биологии рака [4–7]. .Активация Gα i преимущественно связана с ингибированием аденилатциклаз, что приводит к снижению накопления цАМФ в клетках. Однако недавно активация Gα i была связана с несколькими другими молекулярными эффекторами, включая PI3K / Akt [8,9], ERK [10] и c-Src [5].
Измерение активации Gα i обычно проводят путем измерения ингибирования индуцированной форсколином продукции цАМФ. Подобно анализу фосфорилирования дальше по течению, такие измерения не имеют пространственного разрешения, имеют ограниченное временное разрешение и могут зависеть от значительных перекрестных помех и усиления или десенсибилизации сигнала [11–13].
Для исследования активации G-белка прямым путем с высоким пространственно-временным разрешением могут быть использованы генетически закодированные биосенсоры FRET (Förster Resonance Energy Transfer) или BRET (Bioluminescent Resonance Energy Transfer) [14]. Эти методы основаны на измерении безызлучательной передачи энергии от молекулы донора к молекуле акцептора, которая имеет место только тогда, когда донор и акцептор находятся в непосредственной близости друг от друга (<10 нм). Изменения расстояния или ориентации между донорным и акцепторным диполем приводят к изменениям в эффективности RET, которые могут быть определены количественно.
Методы RET позволяют регистрировать кинетику отдельных клеток с миллисекундным разрешением, что может использоваться для определения межклеточной гетерогенности и записи фармакокинетических параметров. Более того, этот подход может регистрировать активацию GPCR в физиологических условиях in vivo [15].
Gα i был успешно помечен люциферазой в различных внутренних сайтах и использован для измерения BRET между различными субъединицами Gα i и GPCR [16–19] или Gγ [19].Также были выполнены измерения FRET между флуоресцентно меченными Gα i1 , Gα i2 и Gα i3 и Gβ [20] или Gγ [21].
Для выполнения измерений FRET необходима спектрально перекрывающаяся пара донора и акцептора [24], и ранее было показано, что использование более ярких флуоресцентных белков может улучшить чувствительность измерений биосенсора FRET [22,23]. Чтобы получить надежные измерения FRET, которые проверяют активацию Gα i , мы соединили Gα i1 Gα i2 и Gα i3 с самым ярким и наиболее фотостабильным мономерным циановым флуоресцентным белком (CFP), доступным в настоящее время, mTurquoise2. (mTq2) [25].В качестве акцептора мы использовали кольцевую пермутированную Венеру (cpVenus), слитую с Gγ 2 , которая ранее использовалась в качестве акцептора в единственном плазмиде Gα q FRET-сенсоре [26]. Мы используем единую плазмидную стратегию для облегчения протоколов трансфекции и обеспечения четко определенного соотношения экспрессии донора и акцептора в клетках [27]. Эта стратегия экспрессии должна значительно облегчить использование и воспроизводимость результатов этих датчиков. Мы представляем стратегию конструирования, проверки и характеристики этого нового поколения датчиков FRET для активации Gα i1 Gα i2 и Gα i3 .Эти биосенсоры очень хорошо подходят для микроскопии живых клеток и могут использоваться для быстрых кинетических измерений в миллисекундном диапазоне, что позволяет определять фармакологические характеристики лекарственного средства и определять on- и off-кинетику для агонистов и антагонистов в Gα и -связанных GPCR.
Мы используем единую плазмидную стратегию для облегчения протоколов трансфекции и обеспечения четко определенного соотношения экспрессии донора и акцептора в клетках [27]. Эта стратегия экспрессии должна значительно облегчить использование и воспроизводимость результатов этих датчиков. Мы представляем стратегию конструирования, проверки и характеристики этого нового поколения датчиков FRET для активации Gα i1 Gα i2 и Gα i3 .Эти биосенсоры очень хорошо подходят для микроскопии живых клеток и могут использоваться для быстрых кинетических измерений в миллисекундном диапазоне, что позволяет определять фармакологические характеристики лекарственного средства и определять on- и off-кинетику для агонистов и антагонистов в Gα и -связанных GPCR.
Результаты
Генерация конструкций
Мономерный вариант CFP mTurquoise2, предпочтительный донор в парах CFP-YFP FRET из-за его высокого квантового выхода и фотостабильности [25], был вставлен в Gα i1 после аланина в положении 121 в петле αB-αC.Этот сайт встраивания, который, как было ранее показано, сохраняет скорости обмена нуклеотидов и GTPase реакции, сопоставимые с белками дикого типа [20]. Gα i1 -mTq2 обнаруживает локализацию плазматической мембраны при экспрессии в клетках HeLa (рис. 1A). Были проведены пробные эксперименты, чтобы проверить, подходит ли Gα i1 -mTq2 для измерения с помощью FRET активации комплекса гетеротримерного G-белка при активации GPCR. С этой целью как Gβ, так и Gγ могут быть помечены акцептором для измерения активации гетеротримерного G-белка с помощью FRET [20,21].Ранее мы показали, что самый высокий контраст FRET для Gα q достигается с cpVenus-Gγ 2 [26]. Поскольку Gα i имеет высокую структурную гомологию с Gα q и сайт, в который вставлен mTurquoise2, аналогичен, мы решили использовать здесь тот же акцептор FRET. Следовательно, чтобы ввести меченый гетеротримерный G-протеиновый комплекс в клетки, мы совместно экспрессировали Gα i1 -mTq2 вместе с акцептором FRET cpVenus-Gγ 2 и немаркированным Gβ 1 [26].
Чтобы изучить коэкспрессию трех субъединиц (Gα i1 -mTq2, Gβ 1 и cpVenus-Gγ 2 ) из одной плазмиды по сравнению с тремя отдельными плазмидами, мы количественно оценили флуоресценцию CFP и YFP в этих двух экспериментальных условия (рис. 1D). Флуоресценция CFP и YFP в стратегии трансфекции одной плазмиды имела коэффициент детерминации r 2 , равный 0.64, тогда как трансфекции тремя отдельными плазмидами показали коэффициент детерминации r 2 0,36 между интенсивностью CFP и интенсивностью YFP. Другими словами, корреляция между экспрессией CFP и YFP лучше в конфигурации с одной плазмидой, что указывает на явное преимущество этой конструкции. Дополнительным преимуществом этой плазмиды является экспрессия белка 3: 1 выше и ниже последовательности IRES, что, как ранее было показано, приводит к предпочтительному соотношению экспрессии донора (CFP) и акцептора (YFP) для аналогичного Gα q FRET. датчик [27].Наконец, отдельные плазмидные конструкции упростят введение в первичные клетки, создание стабильных клеточных линий или трансгенных организмов с Gα i -сенсорами.
Рис. 1. Разработка и характеристика нового датчика Gα i1 .
(A) Типичное изображение, показывающее локализацию на плазматической мембране Gα i1 , слитого с mTurquoise2-Δ9, экспрессированного в клетках HeLa. (B) Схематический обзор плазмиды, содержащей pGβ-2A-YFP-Gγ 2 -IRES-Gα i1 -CFP, управляемую промотором CMV.На вставке показана последовательность ДНК, кодирующая конец последовательности IRES и начало последовательности Gα i1 . Предлагаемая трансляция белка показана в строке под последовательностью ДНК (однобуквенные сокращения аминокислот). (C) Конфокальные изображения локализации Gα i1 -mTurquoise2-Δ9 ( верхний ряд ) и cp173Venus-Gγ 2 ( нижний ряд ) в клетках HeLa, для варианта 1.0 ( левый столбец ) и вариант 2. 0 ( правый столбец ) датчика Gα i1 .(D) Количественный анализ коэкспрессии каналов CFP и YFP трансфекций cp173Venus-Gγ 2 и Gα i1 -mTurquoise2-Δ9 в клетках HeLa. Трансфекция одиночной плазмиды ( слева, ) по сравнению с трансфекцией отдельных плазмид ( справа, ). Точки отображают интенсивность CFP и YFP, количественно определенную для отдельных отдельных ячеек. R 2 — коэффициент детерминации. Ширина отдельных изображений в A и C составляет 143 мкм.
0 ( правый столбец ) датчика Gα i1 .(D) Количественный анализ коэкспрессии каналов CFP и YFP трансфекций cp173Venus-Gγ 2 и Gα i1 -mTurquoise2-Δ9 в клетках HeLa. Трансфекция одиночной плазмиды ( слева, ) по сравнению с трансфекцией отдельных плазмид ( справа, ). Точки отображают интенсивность CFP и YFP, количественно определенную для отдельных отдельных ячеек. R 2 — коэффициент детерминации. Ширина отдельных изображений в A и C составляет 143 мкм.
https: // doi.org / 10.1371 / journal.pone.0146789.g001
Показатели в анализах активации GPCR
Для тестирования нового биосенсора Gα i1 при визуализации живых клеток мы использовали хорошо охарактеризованный GPCR, который, как известно, связан с Gα i1 , адренергическим рецептором α 2 (α 2 AR). Клетки HeLa, которые, как было показано ранее, эндогенно содержат АР α 2 [20], были трансфицированы биосенсором Gα i1 . При добавлении 10 мкМ UK14,304 мы наблюдали устойчивую потерю FRET, измеряя соотношение между флуоресценцией YFP и CFP биосенсора Gα i1 , которое было возвращено к исходному уровню добавлением 60 мкМ α 2 AR. антагонист Йохимбин (рис. 2А). Токсин коклюша (PTX), как было показано, инактивирует передачу сигналов Gα i в клетках посредством АДФ-рибозилирования субъединицы Gα i [28], что предотвращает его взаимодействие с GPCR. Активация Gα i1 была полностью отменена инкубацией в течение ночи с PTX, показывая, что слияние белка Gα i1 -mTq2 все еще остается чувствительным к PTX (рис. 2A). Чтобы подтвердить, что датчик можно использовать для анализа активации эндогенных рецепторов в первичных клетках Gα i1 , мы повторили этот эксперимент на HUVEC (эндотелиальных клетках пупочной вены человека).Добавление хорошо известного стимулятора HUVEC, S1P [29], вызывало стойкое снижение отношения FRET для Gα i1 -сенсора (рис. 2B), ночная обработка PTX полностью устраняла этот ответ. Затем, чтобы исследовать, насколько надежен датчик Gα i1 на других анализах активации GPCR, мы протестировали множество GPCR, которые, как было показано, связаны с Gα i . Рецептор брадикинина 2B (BK 2B ) [30], рецептор лизофосфатидной кислоты 2 (LPA 2 ) [31,32] и рецептор мускаринового ацетилхолина 3 (M 3 ) [33,34] были котрансфицированы биосенсор Gα i1 в клетках HeLa.После стимуляции соответствующими агонистами все три рецептора показали устойчивое снижение соотношения FRET биосенсора Gα i1 (рис. 2C). Рецептор M 3 также показал полное восстановление соотношения FRET до исходного уровня после добавления антагониста атропина. Рецептор M 3 в основном известен своей передачей сигналов через Gα q . Тем не менее, предыдущие исследования показали активацию Gα i через рецептор M 3 [19,33–36], что согласуется с нашими наблюдениями.
2B), ночная обработка PTX полностью устраняла этот ответ. Затем, чтобы исследовать, насколько надежен датчик Gα i1 на других анализах активации GPCR, мы протестировали множество GPCR, которые, как было показано, связаны с Gα i . Рецептор брадикинина 2B (BK 2B ) [30], рецептор лизофосфатидной кислоты 2 (LPA 2 ) [31,32] и рецептор мускаринового ацетилхолина 3 (M 3 ) [33,34] были котрансфицированы биосенсор Gα i1 в клетках HeLa.После стимуляции соответствующими агонистами все три рецептора показали устойчивое снижение соотношения FRET биосенсора Gα i1 (рис. 2C). Рецептор M 3 также показал полное восстановление соотношения FRET до исходного уровня после добавления антагониста атропина. Рецептор M 3 в основном известен своей передачей сигналов через Gα q . Тем не менее, предыдущие исследования показали активацию Gα i через рецептор M 3 [19,33–36], что согласуется с нашими наблюдениями.
Рис. 2. Производительность сенсоров Gα i в анализах передачи сигналов GPCR в одиночных клетках.
(A) Эксперименты по визуализации соотношения FRET в клетках HeLa, трансфицированных датчиком Gα i1 . Быстрая потеря FRET, наблюдаемая по уменьшению отношения YFP / CFP, после стимуляции клеток 10 мкМ UK-14,304, специфического агониста α 2 AR, добавление 60 мкМ йохимбина возвращает соотношение FRET к исходным уровням. Ночная обработка (100 нг / мл) PTX устраняет ответ на Gα i1 -сенсор в клетках, стимулированных UK-14304.(B) Эксперименты по визуализации отношения FRET в Huvecs, трансфицированных датчиком Gα i1 . Устойчивая потеря FRET наблюдается после стимуляции 500 нМ S1P (сфингозин-1-фосфат). Ночная обработка (100 нг / мл) PTX устраняет ответ на Gα i1 -сенсор в S1P-стимулированных клетках. (C) Эксперименты по визуализации соотношения FRET клеток HeLa, трансфицированных датчиком Gα i1 и BK 2B ( вверху справа ), LPA 2 ( вверху слева ), M 3 ( внизу справа ) и β 2 AR-2A2-m Черри ( внизу слева ) стимулировали 1 мкМ брадикинина (BK 2B ), 1 мкМ лизофосфатидовой кислоты (LPA 2 ), 100 мкМ карбахола и 10 мкМ атропина. (M 3 ) или 10 мкМ изопротеренола и 10 мкМ пропранолола (β 2 AR).Клетки HeLa, трансфицированные рецепторами BK 2B , LPA 2 и M 3 , демонстрируют четкое изменение соотношения YFP / CFP FRET при добавлении их соответствующих агонистов, тогда как стимуляция β 2 AR не изменяет FRET. передаточное отношение датчика Gα i1 . В контрольных условиях клетки HeLa, трансфицированные только сенсором Gα i1 , получали идентичные стимуляции. (D) Эксперименты по визуализации соотношения FRET в клетках HeLa, трансфицированных датчиком Gα i2 или датчиком Gα i3 .Быстрая потеря FRET наблюдается после стимуляции клеток 10 мкМ UK-14,304, последующее добавление 60 мкМ йохимбина возвращает соотношение FRET к исходным уровням. Ночная обработка (100 нг / мл) коклюшного токсина (PTX) устраняет ответ на биосенсоры Gα i2 и Gα i3 в клетках, стимулированных UK-14304. Клетки HeLa стимулировали агонистом при t = 32 с, и антагонист добавляли при 90 287 t = 152 с, где указано. Клетки Huvec стимулировали S1P при t = 55 с.Временные диаграммы показывают среднее изменение отношения флуоресценции YFP / CFP (± s.e.m). Средние кривые состоят из данных по меньшей мере 3 независимых экспериментов, проведенных в разные дни, с указанным количеством клеток ( n ) на условие.
(M 3 ) или 10 мкМ изопротеренола и 10 мкМ пропранолола (β 2 AR).Клетки HeLa, трансфицированные рецепторами BK 2B , LPA 2 и M 3 , демонстрируют четкое изменение соотношения YFP / CFP FRET при добавлении их соответствующих агонистов, тогда как стимуляция β 2 AR не изменяет FRET. передаточное отношение датчика Gα i1 . В контрольных условиях клетки HeLa, трансфицированные только сенсором Gα i1 , получали идентичные стимуляции. (D) Эксперименты по визуализации соотношения FRET в клетках HeLa, трансфицированных датчиком Gα i2 или датчиком Gα i3 .Быстрая потеря FRET наблюдается после стимуляции клеток 10 мкМ UK-14,304, последующее добавление 60 мкМ йохимбина возвращает соотношение FRET к исходным уровням. Ночная обработка (100 нг / мл) коклюшного токсина (PTX) устраняет ответ на биосенсоры Gα i2 и Gα i3 в клетках, стимулированных UK-14304. Клетки HeLa стимулировали агонистом при t = 32 с, и антагонист добавляли при 90 287 t = 152 с, где указано. Клетки Huvec стимулировали S1P при t = 55 с.Временные диаграммы показывают среднее изменение отношения флуоресценции YFP / CFP (± s.e.m). Средние кривые состоят из данных по меньшей мере 3 независимых экспериментов, проведенных в разные дни, с указанным количеством клеток ( n ) на условие.
https://doi.org/10.1371/journal.pone.0146789.g002
В условиях контроля, например В отсутствие сверхэкспрессируемого GPCR мы стимулировали клетки HeLa соответствующим агонистом и антагонистом и наблюдали лишь очень незначительный ответ на датчике Gα i1 в случае стимуляции LPA.Скорее всего, это связано с активацией эндогенных рецепторов LPA в клетках HeLa [37]. Когда мы котрансфицировали адренорецептор β 2 (β 2 AR), ни одна из клеток не показала активацию Gα i1 в ответ на лечение агонистом и антагонистом (рис. 2C). Следует отметить, что β 2 AR является классическим активатором Gα s , но при определенных условиях сообщалось о переходе на Gα i [38]. Наши результаты согласуются с единственным известным нам исследованием, в котором используются аналогичные инструменты (сенсоры на основе BRET) и сходные условия (сверхэкспрессия β 2 AR и сенсоры гетеротримерных G-белков) [19].Также в этом случае не наблюдалась активация Gα i1 при стимуляции β 2 AR (и только небольшая активация Gα i2 и Gα i3 , которая была> 10 раз ниже, чем активация альфа-2C. адренорецептор, сильный активатор Gα и ).
2C). Следует отметить, что β 2 AR является классическим активатором Gα s , но при определенных условиях сообщалось о переходе на Gα i [38]. Наши результаты согласуются с единственным известным нам исследованием, в котором используются аналогичные инструменты (сенсоры на основе BRET) и сходные условия (сверхэкспрессия β 2 AR и сенсоры гетеротримерных G-белков) [19].Также в этом случае не наблюдалась активация Gα i1 при стимуляции β 2 AR (и только небольшая активация Gα i2 и Gα i3 , которая была> 10 раз ниже, чем активация альфа-2C. адренорецептор, сильный активатор Gα и ).
Чтобы проверить работоспособность биосенсоров Gα i2 и Gα i3 , мы трансфицировали клетки HeLa их соответствующими плазмидами. Подобно эксперименту с биосенсором Gα i1 на рис. 2А, мы наблюдали устойчивую потерю FRET после добавления 10 мкМ UK14,304, и сигнал вернулся к исходному уровню при добавлении 60 мкМ йохимбина (рис. 2D).В этих экспериментальных условиях мы не наблюдали существенных различий в кинетике активации или амплитуде ответов между тремя различными субъединицами Gα и . И Gα i2 -mTq2, и Gα i3 -mTq2 все еще чувствительны к обработке PTX, как показано отменой ответа FRET после инкубации с PTX в течение ночи (рис. 2D).
Быстрые кинетические измерения
Чтобы более подробно изучить субсекундную кинетику активации Gα i1 в живых клетках, клетки HEK293 были котрансфицированы с помощью Gα i1 -сенсора и α 2 AR или рецептора аденозина A1, соответственно.При использовании системы быстрой перфузии для нанесения лиганда измерения FRET отдельных клеток показывают быструю потерю отношения FRET более чем на 15% после кратковременного применения 20 мкМ норэпинефрина. После отмывания лиганда сигнал FRET возвращается к исходным уровням. Это можно было воспроизвести несколько раз без видимой потери амплитуды сигнала (рис. 3А). Аналогичный ответ наблюдался для аденозинового рецептора A1 после непродолжительного применения эндогенного лиганда аденозина (30 мкМ) (рис. 3B).
3А). Аналогичный ответ наблюдался для аденозинового рецептора A1 после непродолжительного применения эндогенного лиганда аденозина (30 мкМ) (рис. 3B).
Рис 3.Характеристики датчика Gα i1 при кинетических измерениях.
(A) Клетки HEK293, трансфицированные Gα i1 -сенсором и α 2 AR, многократно стимулировали 20 мкМ норадреналина в течение интервалов, обозначенных короткими горизонтальными линиями. Представленные данные являются репрезентативными по крайней мере для шести различных трансфекций, выполненных в течение шести экспериментальных дней. Верхняя панель: эмиссия YFP, центральная панель: эмиссия CFP, нижняя панель: скорректированный и нормализованный коэффициент FRET.(B) Клетки HEK293, трансфицированные Gα i1 -сенсором и аденозиновым A1-рецептором, стимулировали 30 мкМ аденозина, что показано короткой горизонтальной линией. Представленные данные являются репрезентативными по крайней мере для шести различных трансфекций, выполненных в течение шести экспериментальных дней. Верхняя панель: эмиссия YFP, центральная панель: эмиссия CFP, нижняя панель: скорректированный и нормализованный коэффициент FRET. (C) Увеличенное изображение он-кинетики активации Gα i1 , показывающее нормализованное соотношение FRET во время первой стимуляции эксперимента в (A), приспособленное к однокомпонентной экспоненциальной функции затухания с tau = 1160 мс и амплитудой = 0.18 (R = 0,99). (D) Диаграмма разброса, показывающая средние экспоненциальные константы времени (тау) объединенных данных из (n = 10) индивидуальных подборов клеток HEK293, трансфицированных датчиком Gα i1 и α 2 AR, стимулированных 100 мкМ норэпинефрина или объединенных данные (n = 14) по индивидуальным подборам Gα i1 -сенсора и аденозинового A1-рецептора, стимулированного 30 мкМ аденозина, соответственно. Планки погрешностей указывают 95% доверительный интервал.
https://doi.org/10.![]() 1371/journal.pone.0146789.g003
1371/journal.pone.0146789.g003
Эти быстрые измерения FRET можно использовать для оценки кинетики активации Gα i1 с разрешением менее секунды (рис. 3C), как показано крупным планом первой стимуляции в эксперименте, показанном на рис. 3А. Кривая была подогнана к однокомпонентной функции экспоненциального затухания, как описано ранее [39], в результате чего экспоненциальная постоянная времени (τ) составила 1160 мс.
Чтобы оценить точную динамическую кинетику сенсора Gα i1 , клетки стимулировали насыщающими концентрациями лиганда (100 мкМ норэпинефрина или 30 мкМ аденозина).Каждый индивидуальный ответ был подогнан к однокомпонентной экспоненте, что привело к средним значениям τ для α 2 AR, равным 887 мс, и для аденозина A1, равным 963 мс (рис. 3D), что соответствует периодам полувыведения 614 мс и 668 мс соответственно. Эти значения хорошо согласуются с более ранними наблюдениями активации G-белка с помощью FRET [21,40,41].
Заключительные замечания
В этой рукописи мы описываем дизайн, конструкцию и характеристики трех новых биосенсоров FRET для измерения активации Gα i1 , Gα i2 , Gα i3 .Новые сенсоры содержат субъединицу Gα, слитую с донорским флуорофором, mTurquoise2, и субъединицу Gγ, слитую с cp173Venus, поскольку ранее было показано, что эта комбинация для сенсора FRET Gα q обеспечивает наибольший динамический диапазон [26]. Три субъединицы гетеротримеров (Gα i -mTq2, Gβ 1 и cpVenus-Gγ 2 ) были сконфигурированы на одной плазмиде, обеспечивая надежную коэкспрессию с предпочтительной стехиометрией. Мы показываем, что эти датчики хорошо подходят для микроскопии живых клеток и извлечения кинетических параметров с помощью ратиометрической визуализации FRET одной клетки.Стандартизованная компоновка этих биосенсоров FRET для активации G-белка повысит надежность и воспроизводимость экспериментов внутри и между лабораториями. Примером в этой статье является надежная работа датчика Gα i1 в трех разных лабораториях без оптимизации экспериментальных условий.
Примером в этой статье является надежная работа датчика Gα i1 в трех разных лабораториях без оптимизации экспериментальных условий.
Одним из ограничений биосенсоров на основе переноса энергии для гетеротримерных G-белков является то, что они зависят от сверхэкспрессии гетеротримеров, что может влиять на естественное предпочтение GPCR для определенного класса гетеротримерных G-белков.Мечение эндогенных субъединиц флуоресцентными белками потенциально может облегчить это.
Исключительная чувствительность этих датчиков позволяет надежно обнаруживать активацию Gα i в первичных клетках через эндогенные GPCR. Более того, эти биосенсоры можно использовать для прямого сравнения предпочтительных паттернов активации Gα i1 Gα i2 и Gα i3 между различными GPCR, связанными с Gα i , что может помочь в разработке терапевтических стратегий, нацеленных на Gα i . сигнальные пути [42].
Методы
Конструирование флуоресцентных слитых белков
Для вставки mTurquoise2-Δ9 (здесь сокращенно mTq2) [25] в белки Gα i1 , Gα i2 и Gα i3 , версию mTq2 с сайтами рестрикции Age1 на его N-конце и C-конце. был сконструирован путем амплификации mTurquoise2 с прямым праймером 5′-ATaccggttctATGGTGAGCAAGGGCG-3 ‘и обратным праймером 5′-TAaccggtGATCCCGGCGGC-3’. Чтобы ввести Age1-сайт в Gα i1 -цитрин, мы выполнили полновекторную ПЦР на шаблоне RnGalpha i1 -Citrine [20] с прямым праймером 5′-ATaccggtGAACTCGCCGGCGTCATA-3 ‘и обратным праймером 5’-ATaccggtCAGCGCCGTCATAA -3 ‘.Чтобы ввести Age1-сайт в Gα i2 -цитрин, мы выполнили полновекторную ПЦР на матрице HsGalpha i2 -Citrine [20] с прямым праймером 5′-ATaccggtGAGGAGCAAGGCGTGCT-3 ‘и обратным праймером 5’TGTG -3 ‘. КДНК, содержащая кодирующую последовательность для HsGalpha i3 с сайтом Age1, была синтезирована Eurofins (www.eurofins. nl). Разрезание продукта ПЦР mTq2 и новых векторов Gα i1 , Gα i2 и Gα i3 с помощью Age1 и последующее лигирование привело к получению RnGalpha i1 , помеченного mTq2 после позиции 121, и HsGalpha i2,3 , помеченного mTq2 после положения 114, аналогично ранее описанному функционально меченному Gα i1,2,3 [20].
nl). Разрезание продукта ПЦР mTq2 и новых векторов Gα i1 , Gα i2 и Gα i3 с помощью Age1 и последующее лигирование привело к получению RnGalpha i1 , помеченного mTq2 после позиции 121, и HsGalpha i2,3 , помеченного mTq2 после положения 114, аналогично ранее описанному функционально меченному Gα i1,2,3 [20].
Для создания варианта 1.0 сенсора Gα i1 была проведена ПЦР на плазмиде mTq2-Gα i1 с прямым праймером 5′-AGGTCTATATAAGCAGAGC-3 ‘и обратным праймером 5′-TATggatccAGCTTAGAAGAGACCACAGTC-3’ для введения Сайт BamHI на С-конце и сайт NcoI на N-конце. Затем было выполнено тройное лигирование с продуктом ПЦР (разрезанным с помощью BamHI и NcoI), вектором, содержащим pGβ 1 -T2A-cp173Venus-Gγ 2 [43] (разрезанным с помощью BamHI и SacII), и вектором, содержащим pPRIG -IRES [44] (вырезано с помощью NcoI и SacII).Полученная плазмида, pGβ 1 -2A-YFP-Gγ 2 -IRES-MATT-Gα i1 -CFP, коэкспрессирует MATT-Gα i1 -mTurquoise2-Δ9 (нарушение локализации плазматической мембраны), pGβ 1 и cp173Venus-Gγ 2 (рис. 1B).
Чтобы сконструировать вариант 2.0 сенсора Gα i1 , мы выполнили ПЦР с мутагенезом с вариантом 1.0 в качестве матрицы путем амплификации с прямым 5′-GAAAAACACGATGATAATATGGGCTGCACACTGAGC-‘3 и 5’-GCTCAGTGTGCAGCCCATATTATCTTC-3TGT-3.Полученная плазмида, pGβ 1 -2A-cp173Venus-Gγ 2 -IRES-Gα i1 -mTurquoise2-Δ9, коэкспрессирует Gα i1 -mTq2 (правильно расположенный на плазматической мембране), pGβ 1 и cpVenus-Gγ 2 (рис. 1B).
Чтобы создать единый плазмидный сенсор для Gα i2 и Gα i3 , мы выполнили ПЦР с расширением перекрывания [45]. Gα i2 -mTq2 амплифицировали с прямым праймером ‘5-acgatgataatATGGGCTGCACCGTGA-3’ и обратным праймером ‘5 -TATtctagaAGCTCAGAAGAGGCCGCAGT-3’, а Gα i3 -mTq2 амплифицировали с прямым праймером ‘acGGatgata’ 5 праймер ‘5 -TATtctagaAGCTTAATAAAGTCCACATTCCT-3’. Еще одну ПЦР проводили на ранее описанном [26] сенсоре одиночной плазмиды Gα q с прямым праймером «5-GAAGTTTTTCTGTGCCATCC -3» и обратным праймером «5-GCAGCCCATattatcatcgtgtttttcaaag -3». Впоследствии описанный выше продукт ПЦР Gα i2 -mTq2 или Gα i3 -mTq2 смешивали с продуктом ПЦР сенсора Gα q и использовали в качестве матрицы для третьей ПЦР с прямым праймером ‘5-GAAGTTTTTCTGTGCCATCC -3 ‘и обратный праймер’ 5- TATtctagaAGCTCAGAAGAGGCCGCAGT-3 ‘и прямой праймер’ 5- GAAGTTTTTCTGTGCCATCC-3 ‘и обратный праймер’ 5-TATtctagaAGCTTAATAAAGTCCACATTCCT-3 ‘соответственно.Затем полученные продукты ПЦР лигировали в основную цепь сенсора Gα q с помощью SacII и XbaI, в результате чего получали pGβ-2A-cp173Venus -Gγ 2 -IRES-Gα i2 -mTurquoise2-Δ9 и pGβ 1 — 2A-cp173Venus-Gγ2-IRES-Gα i3 -mTurquoise2-Δ9 соответственно. Последовательности плазмид доступны по запросу. Плазмиды будут распространяться через Addgene: http://www.addgene.org/Dorus_Gadella/. RnGα i1 -mCitrine и HsGα i2 -mCitrine были любезным подарком Скотта Гибсона [20].Обратите внимание, что кодирующая последовательность RnGα i1 отличается только одной аминокислотой от Gα i1 человека (S98A). Рецептор LPA 2 был получен с cDNA.org. BK 2 R [46], α 2 AR [21], M 3 R [47] и рецептор A1 [48] были описаны ранее. β 2 AR-P2A-mВишня — подарок от Анны Пьетрашевской (Амстердамский университет).
Еще одну ПЦР проводили на ранее описанном [26] сенсоре одиночной плазмиды Gα q с прямым праймером «5-GAAGTTTTTCTGTGCCATCC -3» и обратным праймером «5-GCAGCCCATattatcatcgtgtttttcaaag -3». Впоследствии описанный выше продукт ПЦР Gα i2 -mTq2 или Gα i3 -mTq2 смешивали с продуктом ПЦР сенсора Gα q и использовали в качестве матрицы для третьей ПЦР с прямым праймером ‘5-GAAGTTTTTCTGTGCCATCC -3 ‘и обратный праймер’ 5- TATtctagaAGCTCAGAAGAGGCCGCAGT-3 ‘и прямой праймер’ 5- GAAGTTTTTCTGTGCCATCC-3 ‘и обратный праймер’ 5-TATtctagaAGCTTAATAAAGTCCACATTCCT-3 ‘соответственно.Затем полученные продукты ПЦР лигировали в основную цепь сенсора Gα q с помощью SacII и XbaI, в результате чего получали pGβ-2A-cp173Venus -Gγ 2 -IRES-Gα i2 -mTurquoise2-Δ9 и pGβ 1 — 2A-cp173Venus-Gγ2-IRES-Gα i3 -mTurquoise2-Δ9 соответственно. Последовательности плазмид доступны по запросу. Плазмиды будут распространяться через Addgene: http://www.addgene.org/Dorus_Gadella/. RnGα i1 -mCitrine и HsGα i2 -mCitrine были любезным подарком Скотта Гибсона [20].Обратите внимание, что кодирующая последовательность RnGα i1 отличается только одной аминокислотой от Gα i1 человека (S98A). Рецептор LPA 2 был получен с cDNA.org. BK 2 R [46], α 2 AR [21], M 3 R [47] и рецептор A1 [48] были описаны ранее. β 2 AR-P2A-mВишня — подарок от Анны Пьетрашевской (Амстердамский университет).
Культура клеток и подготовка проб
клеток HeLa (American Tissue Culture Collection: Manassas, VA, USA) культивировали в Амстердамском университете (Амстердам, Нидерланды) с использованием среды Игла, модифицированной Дульбекко (DMEM), снабженной глутамаксом, 10% FBS, пенициллином (100 Ед / мл). ) и стрептомицин (100 мкг / мл).Все среды для культивирования клеток были получены от Invitrogen (Bleiswijk, NL).
Клетки трансфицировали в 35-миллиметровой чашке со стеклянным покровным стеклом 24 мм Ø # 1 (Menzel-Gläser, Брауншвейг, Германия), используя 1-2 мкл липофектамина 2000 в соответствии с протоколом производителя (Invitrogen), 0,5-1 мкг плазмидной кДНК и 50 мкл OptiMeM (Life Technologies, Блейсвейк, Нидерланды). После инкубации в течение ночи при 37 ° C и 5% CO 2 покровные стекла помещали в камеру для клеток Attofluor (Invitrogen, Breda, NL) и погружали в среду для микроскопии (20 мМ HEPES (PH = 7.4), 137 мМ NaCL, 5,4 мМ KCL, 1,8 мМ CaCL 2 , 0,8 мМ MgCl 2 и 20 мМ глюкозы). Вся микроскопия живых клеток проводилась при 37 ° C.
Эндотелиальные клетки пупочной вены человека (HUVEC) были приобретены у Lonza и культивированы в Sanquin Blood Supply (Амстердам, Нидерланды) на чашках, покрытых FN, в среде EGM-2 с добавлением одиночных клеток (Lonza, Verviers, Бельгия). HUVEC использовали в пассаже номер 4 или 5. В качестве метода трансфекции использовали систему трансфекции Neon (MPK5000, Invitrogen) и соответствующий набор для трансфекции Neon (Invitrogen).Один импульс генерировали при 1300 В в течение 30 мс для микропорации HUVEC с 2 мкг кДНК, затем клетки высевали на покровные стекла, покрытые FN.
Для быстрых кинетических измерений активации Gα i1 клетки HEK293 культивировали в Университете Вюрцбурга (Вюрцбург, Германия) в среде Игла, модифицированной Дульбекко (DMEM) с добавлением 10% фетальной телячьей сыворотки, L-глутамина (2 мМ). (PAN Biotech GmbH, Айденбах, Германия), пенициллин (100 Ед / мл) и стрептомицин (100 мкг / мл) и хранили при 37 ° C в атмосфере 7% CO2.Клетки собирали и высевали на 24-миллиметровые покровные стекла, покрытые D-полилизином, при ~ 40% конфлюентности. Через три часа клетки временно трансфицировали 1,0 мкг рецептора (α 2 AR или аденозин A1) и 3,0 мкг pGβ 1 -2A-YFP-Gγ 2 -IRES-Gα i1 -mTq2 кДНК на 6 -луночный планшет с использованием реагента для трансфекции Effectene ® (Qiagen) в соответствии с протоколом производителя. Среду для выращивания обновляли через 24 часа, и измерения проводили после общего времени инкубации 48 часов.Клетки содержали в среде для микроскопии (140 мМ NaCl, 5,4 мМ KCl, 2 мМ CaCl2, 1 мМ MgCl2, 10 мМ HEPES, pH 7,3) и постоянно суперслили с этим буфером или буфером, дополненным соответствующим лигандом, с использованием компьютерной системы. Устройство быстрой суперфузии с электромагнитным клапаном (ValveLink 8.2, Automate Scientific).
Среду для выращивания обновляли через 24 часа, и измерения проводили после общего времени инкубации 48 часов.Клетки содержали в среде для микроскопии (140 мМ NaCl, 5,4 мМ KCl, 2 мМ CaCl2, 1 мМ MgCl2, 10 мМ HEPES, pH 7,3) и постоянно суперслили с этим буфером или буфером, дополненным соответствующим лигандом, с использованием компьютерной системы. Устройство быстрой суперфузии с электромагнитным клапаном (ValveLink 8.2, Automate Scientific).
Широкопольная микроскопия
Ратиометрические измерения FRET в клетках HeLa (результаты представлены на рис. 2A, 2C и 2D) были выполнены с использованием широкопольного флуоресцентного микроскопа (Axiovert 200 M; Carl Zeiss GmbH, Германия) в Амстердамском университете (Амстердам, Нидерланды). выдерживали при 37 ° C, снабжали иммерсионным масляным объективом (Plan-Neoflor 40 × / 1.30; Carl Zeiss GmbH) и ксеноновую дуговую лампу с монохроматором (Cairn Research, Faversham, Kent, UK). Изображения были записаны с помощью охлаждаемой камеры устройства с заряженной связью (Coolsnap HQ, Roper Scientific, Tucson, AZ, США). Типичное время экспозиции составляло от 75 мс до 150 мс, а интервал камеры был установлен на 4×4. Флуорофоры возбуждались светом 420 нм (ширина щели 30 нм) и отражались на образец дихроичным зеркалом 455DCLP, эмиссия CFP регистрировалась фильтром BP470 / 30, а эмиссия YFP регистрировалась фильтром BP535 / 30 путем вращения колеса фильтров. .В экспериментах по совместной экспрессии YFP возбуждали светом 500 нм (ширина щели 30 нм) и отражали на образец дихроичным 515DCXR, а излучение регистрировали с помощью фильтра BP535 / 30. Регистрация была скорректирована на фоновый сигнал и, для изображения отношения FRET, просачивание эмиссии CFP в канал YFP (55% интенсивности, измеренной в канале CFP).
Для экспериментов FRET в HUVEC (результаты представлены на рис. 2B) микроскоп Zeiss Observer Z1 использовался в Sanquin Blood Supply (Амстердам, Нидерланды) с 40x NA 1.3 масляный иммерсионный объектив и возбуждающий источник света HXP 120 В. CFP возбуждали через кубический фильтр FRET (Exciter ET 436 / 20x и дихроичное зеркало 455 DCLP (Chroma, Bellows Falls, Вермонт, США)). Излучение направлялось на прикрепленный адаптер двойной камеры (Carl Zeiss GmbH, Германия), управляющий дихроичным зеркалом 510 DCSP (Chroma, Bellows Falls, Вермонт, США). Эмиссионные волны с длиной волны 455–510 нм направляются на эмиссионный фильтр ET 480/40 (Chroma, Bellows Falls, Вермонт, США), а затем фиксируются камерой Hamamatsu ORCA-R2.Длина волны излучения 510 нм и выше направляется на эмиссионный фильтр ET 540/40 м (Ludl Electronic Products, Нью-Йорк, США), а затем фиксируется второй камерой Hamamatsu ORCA-R2. Получение изображений производилось с использованием программного обеспечения микроскопа Zeiss-Zen 2011. Все измерения были скорректированы на фоновый сигнал. Регистрация была скорректирована на фоновый сигнал и утечку эмиссии CFP в канале YFP (62% от интенсивности, измеренной в канале CFP).
CFP возбуждали через кубический фильтр FRET (Exciter ET 436 / 20x и дихроичное зеркало 455 DCLP (Chroma, Bellows Falls, Вермонт, США)). Излучение направлялось на прикрепленный адаптер двойной камеры (Carl Zeiss GmbH, Германия), управляющий дихроичным зеркалом 510 DCSP (Chroma, Bellows Falls, Вермонт, США). Эмиссионные волны с длиной волны 455–510 нм направляются на эмиссионный фильтр ET 480/40 (Chroma, Bellows Falls, Вермонт, США), а затем фиксируются камерой Hamamatsu ORCA-R2.Длина волны излучения 510 нм и выше направляется на эмиссионный фильтр ET 540/40 м (Ludl Electronic Products, Нью-Йорк, США), а затем фиксируется второй камерой Hamamatsu ORCA-R2. Получение изображений производилось с использованием программного обеспечения микроскопа Zeiss-Zen 2011. Все измерения были скорректированы на фоновый сигнал. Регистрация была скорректирована на фоновый сигнал и утечку эмиссии CFP в канале YFP (62% от интенсивности, измеренной в канале CFP).
Для быстрых кинетических измерений активации Gα i1 (результаты представлены на рис. 3) визуализацию выполняли на инвертированном микроскопе Zeiss Axiovert 200 в Университете Вюрцбурга (Вюрцбург, Германия), оборудованном иммерсионным объективом 63x и фотометрическая система с двойным излучением (Till Photonics), описанная ранее [21].Трансфицированные клетки возбуждали светом от полихрома IV (Till Photonics). Освещение было установлено на 40 мс из общего времени интеграции 100 мс. Сигналы CFP (480 ± 20 нм), YFP (535 ± 15 нм) и отношения FRET (YFP / CFP) регистрировались одновременно (светоделитель DCLP 505 нм) при возбуждении на длине волны 436 ± 10 нм (светоделитель DCLP 460 нм). Сигналы флуоресценции регистрировались фотодиодами и оцифровывались с помощью аналого-цифрового преобразователя (Digidata 1440A, Axon Instruments). Все данные были записаны на ПК под управлением Clampex 10.3 (Axon Instruments). Полученные отдельные трассы были подогнаны к однокомпонентной функции экспоненциального затухания для извлечения экспоненциальной постоянной времени tau [39]. Полупериод активации (t 1/2 ) определяется как τ * ln2. В динамических экспериментах клетки стимулировали UK14,304 (10 мкм), йохимбином (60 мкм), брадикинином (1 мкм), LPA (1 мкм), карбахолом (100 мкм), атропином (10 мкм), изопротеренолом (10 мкм), пропранололом (10 мкм), S1P (500 нМ), 20 мкМ или 100 мкМ норэпинефрина или 30 мкМ аденозина в указанные моменты времени.ImageJ (Национальный институт здоровья) использовался для анализа необработанных изображений микроскопии. Дальнейшая обработка данных была проведена в Excel (Microsoft Office), а графики и статистика были выполнены с использованием Graphpad версии 6.0 для Mac, GraphPad Software, La Jolla California USA, www.graphpad.com.
Полупериод активации (t 1/2 ) определяется как τ * ln2. В динамических экспериментах клетки стимулировали UK14,304 (10 мкм), йохимбином (60 мкм), брадикинином (1 мкм), LPA (1 мкм), карбахолом (100 мкм), атропином (10 мкм), изопротеренолом (10 мкм), пропранололом (10 мкм), S1P (500 нМ), 20 мкМ или 100 мкМ норэпинефрина или 30 мкМ аденозина в указанные моменты времени.ImageJ (Национальный институт здоровья) использовался для анализа необработанных изображений микроскопии. Дальнейшая обработка данных была проведена в Excel (Microsoft Office), а графики и статистика были выполнены с использованием Graphpad версии 6.0 для Mac, GraphPad Software, La Jolla California USA, www.graphpad.com.
Конфокальная микроскопия
клеток HeLa, трансфицированных указанными конструкциями, получали с помощью конфокального микроскопа Nikon A1, оснащенного иммерсионным объективом 60x (Plan Apochromat VC, NA 1.4). Размер точечного отверстия был установлен на 1 единицу Эйри (<0,8 мкм).
Образцы последовательно возбуждали лазерной линией 457 нм и 514 нм и отражались на образец дихроичным зеркалом 457/514. Эмиссия CFP фильтровалась через эмиссионный фильтр BP482 / 35; Эмиссию YFP фильтровали через эмиссионный фильтр BP540 / 30. Чтобы избежать проступания, изображения были получены в режиме последовательной строчной развертки. Все измерения были скорректированы на фоновый сигнал.
Благодарности
Мы благодарим Eelco Hoogendoorn (Амстердамский университет) и Marten Postma (Амстердамский университет) за помощь в анализе данных.Мы благодарим Jorrit Broertjes (студент-магистр Амстердамского университета) и Денниса Ботмана (магистр наук в Амстердамском университете) за создание Gα i1 -mTurquoise2 и Gα i2 -mTurquoise2 соответственно. Мы благодарим Мерел Аджобо-Херманс, Анну Пьетрашевскую и Скотта Гибсона за предоставленные плазмиды.
Вклад авторов
Задумал и спроектировал эксперименты: JvU PLH CH JG TWG. Проведены эксперименты: JvU ADS BS NRR. Проанализированы данные: JvU ADS BS NRR CH JG.Написал статью: JvU ADS BS NRR PLH CH TWG JG.
Проведены эксперименты: JvU ADS BS NRR. Проанализированы данные: JvU ADS BS NRR CH JG.Написал статью: JvU ADS BS NRR PLH CH TWG JG.
Ссылки
- 1. Суки В.Н., Абрамовиц Дж., Маттера Р., Кодина Дж., Бирнбаумер Л. Геном человека кодирует по меньшей мере три неаллеллических G-белка с субъединицами альфа i-типа. FEBS Lett. 1987; 220: 187–192. pmid: 2440724
- 2. Kimple ME, Neuman JC, Linnemann AK, Casey PJ. Ингибирующие G-белки и их рецепторы: новые терапевтические мишени при ожирении и диабете. Exp Mol Med. 2014; 46: e102. pmid: 24946790
- 3.Ван И, Ли Й, Ши Г. Регулирующая функция гетеротримерных белков G в иммунной системе. Arch Immunol Ther Exp (Warsz). 2013; 61: 309–319.
- 4. Дорсам РТ, Гуткинд Ж.С. Рецепторы, сопряженные с G-белком, и рак. Nat Rev Рак. 2007; 7: 79–94. pmid: 17251915
- 5. Даака Ю. Белки G при раке: парадигма рака простаты. Sci STKE. 2004; 2004: re2. pmid: 14734786
- 6. Appleton KM, Bigham KJ, Lindsey CC, Hazard S, Lirjoni J, Parnham S и др.Разработка ингибиторов гетеротримерных субъединиц Gαi. Bioorg Med Chem. 2014; 22: 3423–3434. pmid: 24818958
- 7. Раймонд Дж. Р., Эпплтон К. М., Пирс Дж. Ю., Петерсон Ю. К.. Подавление сообщения GNAI2 при раке яичников. J Ovarian Res. 2014; 7: 6. pmid: 24423449
- 8. Гарсия А., Ким С., Бхавараджу К., Шенвалдер С.М., Кунапули С.П. Роль фосфоинозитид-3-киназы бета в агрегации тромбоцитов и генерации тромбоксана А2, опосредованной сигнальными путями Gi. Биохим Дж.2010. 429: 369–377. pmid: 20441566
- 9. la Sala A, Gadina M, Kelsall BL. G (i) -зависимое ингибирование продукции IL-12 опосредуется активацией пути фосфатидилинозитол-3-киназа-протеин-3-киназа B / Akt и JNK. J Immunol. 2005; 175: 2994–2999. pmid: 16116186
- 10. Kue PF, Daaka Y. Существенная роль G-белков в росте клеток рака простаты и передаче сигналов. J Urol. 2000. 164: 2162–2167. pmid: 11061948
- 11. Hur EM, Kim KT.Передача сигналов рецептора, связанного с G-белком, и перекрестные помехи: достижение быстроты и специфичности. Сотовый сигнал. 2002; 14: 397–405. pmid: 11882384
- 12. Брюс ДжиЭ, Штрауб С.В., Йол Д.И. Перекрестные помехи между передачей сигналов цАМФ и Са2 + в невозбудимых клетках. Клеточный кальций. 2003. 34: 431–444. pmid: 14572802
- 13. Фей Д., Краучер Д. Р., Колч В., Холоденко Б. Н.. Перекрестные помехи и переключатели сигналов в каскадах митоген-активируемых протеинкиназ. Front Physiol. 2012; 3: 355. pmid: 23060802
- 14.Lohse MJ, Nuber S, Hoffmann C. Методы резонансной передачи энергии флуоресценции / биолюминесценции для изучения активации и передачи сигналов рецепторов, связанных с G-белками. Pharmacol Rev.2012; 64: 299–336. pmid: 22407612
- 15. ван Унен Дж., Вулард Дж., Ринкен А., Хоффманн С., Хилл С., Годхарт Дж. и др. Перспектива изучения передачи сигналов GPCR с помощью RET-биосенсоров в живых организмах. Mol Pharmacol. 2015. 88 (3): 589–595.
- 16. Гейлс С., Ван Дурм Дж.Дж., Шаак С., Понтье С., Першрансье Й., Одет М. и др.Исследование структурных перестроек, вызванных активацией, в предварительно собранных комплексах рецептор-G-белок. Nat Struct Mol Biol. 2006; 13: 778–786. pmid: 168
- 17. Gales C, Rebois RV, Hogue M, Trieu P, Breit A, Hébert TE и др. Мониторинг в реальном времени взаимодействий рецепторов и G-белков в живых клетках. Нат методы. 2005. 2: 177–184. pmid: 15782186
- 18. Ayoub MA, Maurel D, Binet V, Fink M, Prezeau L, Ansanay H, et al. Анализ в реальном времени вызванной агонистами активации протеазо-активированного комплекса рецептор 1 / Galphai1, измеренный с помощью биолюминесцентного резонансного переноса энергии в живых клетках.Mol Pharmacol. 2007. 71: 1329–1340. pmid: 17267663
- 19. Saulière A, Bellot M, Paris H, Denis C, Finana F, Hansen JT и др. Расшифровка сложности предвзятого агонизма выявляет новый активный объект рецептора AT1. Nat Chem Biol. Издательская группа «Природа»; 2012; 8: 623–631.
- 20. Гибсон СК, Гилман АГ. Субъединицы Giα и Gβ определяют селективность активации G-белка α2-адренорецепторами. Proc Natl Acad Sci USA. Национальная академия наук; 2006; 103: 212.
- 21. Бюнеманн М., Франк М., Лозе М.Дж. С обложки: Активация белка Gi в интактных клетках связана с перестройкой субъединиц, а не с диссоциацией. Proc Natl Acad Sci USA. Национальная академия наук; 2003; 100: 16077.
- 22. Klarenbeek JB, Goedhart J, Hink MA, Gadella TWJ, Jalink K. Датчик цАМФ на основе mTurquoise как для FLIM, так и для логометрического считывания имеет улучшенный динамический диапазон. PLoS ONE. 2011; 6: e19170. pmid: 21559477
- 23. Спарта Б, Паргетт М, Минге М, Дистор К, Белл Дж, Олбек Дж.Для импульсов активности киназы, регулируемой внеклеточным сигналом (ERK), необходимы механизмы рецепторного уровня. J Biol Chem. 2015; 290: 24784–24792. pmid: 26304118
- 24. Hamers D, van Voorst Vader L, Borst JW, Goedhart J. Разработка биосенсоров FRET для систем млекопитающих и растений. Протоплазма. 2014; 251: 333–347. pmid: 24337770
- 25. Goedhart J, Stetten von D, Noirclerc-Savoye M, Lelimousin M, Joosen L, Hink MA и др.Структурно-управляемая эволюция голубых флуоресцентных белков в направлении квантового выхода 93%. Nature Communications. 2012; 3: 751. pmid: 22434194
- 26. Adjobo-Hermans MJW, Goedhart J, van Weeren L, Nijmeijer S, Manders EMM, Offermanns S, et al. Визуализация в реальном времени активации гетеротримерного G-белка Gq в живых клетках. BMC Biol. 2011; 9: 32. pmid: 21619590
- 27. Goedhart J, van Weeren L, Adjobo-Hermans MJW, Elzenaar I, Hink MA, Gadella TWJ. Количественная совместная экспрессия белков на уровне отдельной клетки — приложение к мультимерному датчику FRET.Дельпрато А.М., редактор. PLoS ONE. 2011; 6: e27321. pmid: 22114669
- 28. Бернс DL. Субъединичная структура и ферментативная активность токсина коклюша. Microbiol Sci. 1988. 5: 285–287. pmid: 28
- 29. Ван Ф., Ван Броклин Дж. Р., Хобсон Дж. П., Мовафаг С., Зуковска-Гройец З., Мильстиен С. и др. Сфингозин-1-фосфат стимулирует миграцию клеток через рецептор клеточной поверхности, связанный с G (i). Возможное участие в ангиогенезе. J Biol Chem. 1999; 274: 35343–35350. pmid: 10585401
- 30.Ляо JK, Homcy CJ. Белки G семейства G alpha i и G alpha q связывают рецептор брадикинина с высвобождением релаксирующего фактора эндотелия. J Clin Invest. 1993; 92: 2168–2172. pmid: 8227332
- 31. ван Корвен EJ, Groenink A, Jalink K, Eichholtz T., Moolenaar WH. Пролиферация клеток, индуцированная лизофосфатидатом: идентификация и анализ сигнальных путей, опосредованных G-белками. Клетка. 1989; 59: 45–54. pmid: 2551506
- 32. An S, Bleu T, Hallmark OG, Goetzl EJ.Характеристика нового подтипа человеческого рецептора, связанного с G-белком, для лизофосфатидной кислоты. J Biol Chem. 1998; 273: 7906–7910. pmid: 9525886
- 33. Offermanns S, Wieland T, Homann D, Sandmann J, Bombien E, Spicher K и др. Трансфицированные мускариновые рецепторы ацетилхолина селективно связываются с белками G Gi-типа и Gq / 11. Mol Pharmacol. 1994; 45: 890–898. pmid: 81
- 34. Akam EC, Challiss RA, Nahorski SR. Профили активации G (q / 11) и G (i / o) в клетках СНО, экспрессирующих человеческие мускариновые рецепторы ацетилхолина: зависимость от агониста, а также от подтипа рецептора.Br J Pharmacol. 2001; 132: 950–958. pmid: 11181437
- 35. Хорниголд, округ Колумбия, Мистри Р., Раймонд П.Д., Бланк Д.Л., Челлисс Р.Дж. Доказательства перекрестного взаимодействия между мускариновыми ацетилхолиновыми рецепторами М2 и М3 в регуляции вторичных мессенджеров и сигнальных путей киназ, регулируемых внеклеточными сигналами, в клетках яичников китайского хомячка. Br J Pharmacol. 2003. 138: 1340–1350. pmid: 12711635
- 36. Blaukat A, Barac A, Cross MJ, Offermanns S, Dikic I. G-белок-связанный рецептор-опосредованная активация митоген-активируемой протеинкиназы через взаимодействие сигналов Galpha (q) и Galpha (i).Молекулярная и клеточная биология. 2000. 20: 6837–6848. pmid: 10958680
- 37. Jongsma M, Matas-Rico E, Rzadkowski A, Jalink K, Moolenaar WH. LPA — хеморепеллент для клеток меланомы B16: действие через рецептор LPA5, повышающий цАМФ. PLoS ONE. 2011; 6: e29260. pmid: 22195035
- 38. Хилл SJ, Бейкер JG. Взлеты и падения переключения Gs- на Gi-белок. Br J Pharmacol. 2003; 138: 1188–1189. pmid: 12711616
- 39. Hoffmann C, Gaietta G, Bünemann M, Adams SR, Oberdorff-Maass S, Behr B и др.Подход FRET на основе FlAsH для определения активации рецепторов, связанных с G-белками, в живых клетках. Нат методы. 2005; 2: 171–176. pmid: 15782185
- 40. Хайн П., Роша Ф., Хоффманн С., Дорш С., Николаев В.О., Энгельхардт С. и др. Активация GS ограничена по времени при инициировании передачи сигналов, опосредованной рецептором. Журнал биологической химии. 2006; 281: 33345–33351. pmid: 16963443
- 41. Hoffmann C, Nuber S, Zabel U, Ziegler N, Winkler C, Hein P и др. Сравнение кинетики активации M3-ACh-рецептора и конститутивно активного мутантного рецептора в живых клетках.Mol Pharmacol. 2012; 82 (2): 236–245 pmid: 22564786
- 42. Деванатан В., Хагедорн И., Келер Д., Пекса К., Черпокова Д., Крафт П. и др. Белок Gαi2 тромбоцитов является важным медиатором тромбовоспалительного поражения органов у мышей. Proc Natl Acad Sci USA. 2015; 112: 6491–6496. pmid: 25944935
- 43. Нагаи Т., Ямада С., Томинага Т., Итикава М., Мияваки А. Расширенный динамический диапазон флуоресцентных индикаторов для Ca (2+) за счет циркулярно пермутированных желтых флуоресцентных белков.Proc Natl Acad Sci USA. 2004; 101: 10554–10559. pmid: 15247428
- 44. Мартин П., Албагли О., Погги М.С., Булукос К.Э., Погнонец П. Разработка нового бицистронного ретровирусного вектора с сильной активностью IRES. BMC Biotechnol. 2006; 6: 4. pmid: 16409632
- 45. Хекман К.Л., Пиз Л.Р. Сплайсинг генов и мутагенез с помощью ПЦР-управляемого удлинения перекрытия. Nat Protoc. 2007; 2: 924–932. pmid: 17446874
- 46. Adjobo-Hermans MJW, Goedhart J, Gadella TWJ.Регулирование заякоривания мембраны PLCbeta1a его субстратом фосфатидилинозитол (4,5) -бисфосфатом. Журнал клеточной науки. 2008; 121: 3770–3777. pmid: 18957514
- 47. Ziegler N, Bätz J, Zabel U, Lohse MJ, Hoffmann C. Датчики на основе FRET для человеческих рецепторов M1-, M3- и M5-ацетилхолина. Bioorg Med Chem. 2011; 19: 1048–1054. pmid: 20716489
- 48. Klotz KN, Hessling J, Hegler J, Owman C, Kull B., Fredholm BB, et al. Сравнительная фармакология подтипов аденозиновых рецепторов человека — характеристика стабильно трансфицированных рецепторов в клетках СНО.Naunyn Schmiedebergs Arch Pharmacol. 1998; 357: 1–9. pmid: 9459566
Гриф гитары | Интернет-галерея Autodesk
© Autodesk, Inc., 2014. Все права защищены.
Использование данного Сервиса регулируется условиями применимых условий обслуживания Autodesk, принятых при доступе к нему.
Эта Служба может включать или использовать фоновые компоненты технологии Autodesk. Для получения информации об этих компонентах щелкните здесь: http: // www.autodesk.com/cloud-platform-components
Товарные знаки
Autodesk, логотип Autodesk и Fusion 360 являются зарегистрированными товарными знаками или товарными знаками Autodesk, Inc. и / или ее дочерних и / или дочерних компаний.
Все остальные торговые марки, названия продуктов или товарные знаки принадлежат их соответствующим владельцам.
Авторские права и авторство стороннего программного обеспечения
Рубиновые драгоценные камни являются Авторскими правами (c) Чад Фаулер, Рич Килмер, Джим Вейрих и другие. Авторские права на части (c) Engine Yard и Andre Arko
bootstrap-select.js — Copyright (C) 2013 bootstrap-select
Backbone.js — Copyright (c) 2010-2013 Джереми Ашкенас, DocumentCloud
Apple-Style Flip Counter — Copyright (c) 2010 Chris Nanney
imagesLoaded © 2013 David DeSandro
jQuery — Copyright 2013 jQuery Foundation и другие участники http://jquery.com/
jQuery timepicker addon — Copyright (c) 2013 Трент Ричардсон
jQuery ColorBox — Copyright (c) 2013 Jack Moore
jQuery.Авторские права принадлежат gritter (c) 2013 Jordan Boesch
Masonry Copyright (c) 2013 David DeSandro
Underscore — Copyright (c) 2009-2013 Jeremy Ashkenas, DocumentCloud and Investigative
Reporters & Editors
underscore_string является Copyright (c) 2011 Esa-Matti Suuronen [email protected]
Icanhaz.js — это ICanHaz.js — Copyright (c) Хенрик Йоретег, 2010 (Mustache и Mustache.js — Copyright (c) 2009 Chris Wanstrath (Ruby) и Copyright (c) ) 2010 Ян Ленардт (JavaScript) соответственно)
Calendario является Авторским правом (c) Codrops 2014, tympanus
Все вышеперечисленные программные компоненты находятся под лицензией MIT.
Настоящим предоставляется бесплатное разрешение любому лицу, получившему копию этого программного обеспечения и связанных файлов документации («Программное обеспечение»), на использование Программного обеспечения без ограничений, включая, помимо прочего, права на использование, копирование, изменение , объединять, публиковать, распространять, сублицензировать и / или продавать копии Программного обеспечения и разрешать лицам, которым предоставляется Программное обеспечение, делать это при соблюдении следующих условий:
Приведенное выше уведомление об авторских правах и это уведомление о разрешении должны быть включены во все копии или существенные части Программного обеспечения.
ПРОГРАММНОЕ ОБЕСПЕЧЕНИЕ ПРЕДОСТАВЛЯЕТСЯ «КАК ЕСТЬ», БЕЗ КАКИХ-ЛИБО ГАРАНТИЙ, ЯВНЫХ ИЛИ ПОДРАЗУМЕВАЕМЫХ, ВКЛЮЧАЯ, НО НЕ ОГРАНИЧИВАЯСЬ ГАРАНТИЯМИ КОММЕРЧЕСКОЙ ЦЕННОСТИ, ПРИГОДНОСТИ ДЛЯ ОПРЕДЕЛЕННЫХ ЦЕЛЕЙ И НЕ ЗАЩИЩАЕТСЯ. НИ ПРИ КАКИХ ОБСТОЯТЕЛЬСТВАХ АВТОРЫ ИЛИ ВЛАДЕЛЬЦЫ АВТОРСКИХ ПРАВ НЕ НЕСЕТ ОТВЕТСТВЕННОСТИ ЗА ЛЮБЫЕ ПРЕТЕНЗИИ, УБЫТКИ ИЛИ ДРУГИЕ ОТВЕТСТВЕННОСТЬ, БЫЛИ В РЕЗУЛЬТАТЕ ДОГОВОРА, ПРАКТИЧЕСКИХ ПРАВ ИЛИ ИНЫХ СЛУЧАЕВ, ВЫЯВЛЕННЫХ, ВНУТРИ ИЛИ В СВЯЗИ С ПРОГРАММНЫМ ОБЕСПЕЧЕНИЕМ ИЛИ ИСПОЛЬЗОВАНИЕМ ИЛИ ДРУГИМИ ДЕЛАМИ ПРОГРАММНОЕ ОБЕСПЕЧЕНИЕ.
На части, относящиеся к лайтбоксу, распространяется лицензия Creative Commons Attribution 2.5 Лицензия (http://creativecommons.org/licenses/by/2.5/). Автором лайтбокса является Локеш Дхакар (lokeshdhakar.com).
клей на гриф Yinfente гитары 22 лад 24,75 дюйма красное дерево палисандр Клей для грифа на LP электрогитаре инкрустация с птицами необработанная
клей на гриф Yinfente гитаре 22 лад 24,75 дюйма красное дерево палисандр Клей на гриф электрогитары с птицей незавершенный
инкрустация для гитары лад 24,75-дюймовый клей для грифа из красного дерева и палисандра на электрогитаре LP Необработанная инкрустация с птицами (наклеивается): Музыкальные инструменты.Купить Гриф Yinfente Гриф 22 лад 24,75 дюйма Гриф из красного дерева палисандр Клей для электрогитары LP Необработанная инкрустация с птицами (наклеивание): Шеи — ✓ БЕСПЛАТНАЯ ДОСТАВКА при покупке, отвечающей критериям. Гриф электрогитары New Mahogany Wood Гриф из розового дерева Инкрустация с птицами 22 лада 24,75 дюйма。 Анкерный стержень на грифе, крепление грифа, нормальная гайка, ширина гайки 42 мм 。 Список пакетов: 1 * гриф гитары。 Гриф электрогитары марки Yinfente. Все сделано из массива дерева. Много лет мастерил гитарой.С высококачественным деревянным материалом. Эта гитарная шея предназначена для замены гитарной шеи в стиле LP. Вы можете установить его на корпусе гитары. 。。。
клей на грифе гитары Yinfente 22 лад 24,75 дюйма красное дерево палисандр Наклейка грифа на электрогитаре LP инкрустация птиц незаконченная
Молоток Yamaha KPB-91B. Yamaha PKBZ1 Регулируемая подставка для клавиатуры в стиле Z, комплект MK-SD / PLBURST с тюнером Kala и полировальной тканью Lumintrail Kala Purple Burst Dolphin Makala Soprano Ukulele.# 5 Easy Stick Twirl… Подходит для начинающих барабанщиков. Аксессуары для барабанных палочек Захват или зажимы для управления. Принадлежность США Замените зарядное устройство с адаптером постоянного тока 9 В переменного тока для кабеля питания клавиатуры CTK-4000 CTK-558. Dean GS B6 TSB Gran Sport 6-String Solid Body Banjo Tobacco Sunburst, Evans Clear 200 Боковая головка малого барабана 13 дюймов. Новый адаптер переменного / постоянного тока для Casio CZ-101 MA-170 GZ-500 GZ-50M CZ101 MA170 GZ500 GZ50M Кабель питания для синтезатора клавиатуры PS Wall Home Charger Worldwide. Пара Mackie CR4 4 Creative Reference Мультимедийные мониторы Колонки + подставки + колодки, домашние тапочки с динозаврами для девочек и мальчиков.FURUTECH iHP-35M 1,3-метровые аудионаушники с возможностью записи, 36-дюймовые стойки динамиков для студийных мониторов Rockville для мониторов Yamaha HS5. ETP-UV2-R Evans UV2 с покрытием Tom Pack Rock 10, 12, 16, Topeakmart Duet Piano Bench Piano с мягкой подушкой и сиденьем для клавиатуры для хранения музыки Коричневый. G-SHOCK-8L Gator Cases Легкий литой противоударный кейс с глубиной 16,5 мм; Размер 8U. Глянцевые белые пластики Aquarian SKPII18WH Super-Kick II Prepack 18-дюймовая пластина для бас-барабана. Цифровые пианино и консоли Защитный пылезащитный чехол для клавиатуры 61 Защитный чехол для клавиатуры цвета хаки, комплект для монтажа в стойку Avaya IPO.NIHAI DIY 5D Diamond Painting Kit Сова Специальная форма Сверло Алмазная вышивка Картины Стразы Картина Вышивка крестиком Искусство Ремесло Холст для гостиной Спальня Home Decor.
Высота действия Gretsch
Высота струны (действие) напрямую связана с интонацией. Эффекты повышения и понижения высоты струны Мне нравится, чтобы высота струны на первом ладу была такой же или немного меньшей, как когда я ладаю на первом ладу и.Таблица размеров рамы велосипеда в дюймах в сантиметрах
Спрячь дев и молись! Впервые появившись здесь, на FigureRealm, я с особым удовольствием представляю … The Cramps! Американская рок-группа, основанная в 1976 году, The Cramps испытала сильное влияние раннего рокабилли, ритм-н-блюза и рок-н-ролльных риффов.Смешивание горячего пуха битника из космоса с хоррорами из B-фильмов, вокалистом Lux Interior и горячим гитаристом тамале Poison Ivy Watch house, сезон 1
Регулировка высоты гитарного гайки поможет вам отрегулировать действие вашего инструмента. В этой статье мы рассмотрим правильную высоту затяжки гайки, чтобы вы могли правильно настроить гитару без … Кейсы для мобильных криминалистов
Я лично профессионально настрою каждый инструмент (угол грифа, рельеф грифа, действие струн, интонацию, высоту звукоснимателя и т. Д.), Включая новый набор струн с калибром струн по вашему выбору. Я отправлю все, что угодно, в любую точку мира, в надежной упаковке — сразу после получения вашего платежа. Fdr первый депозит данных pnc
Высота. 6 футов 3½ дюйма или 191,5 см. Масса. 95 кг или 209,5 фунтов. Подруга / супруга. Стив Хоуи встречался с Сарой Шахи (2004-настоящее время) — Стив начал встречаться с актрисой Сарой Шахи в августе 2004 года. В конце концов, во время отпуска на Гавайях Стив сделал предложение Саре, и в июне 2007 года они обручились, а позже связали себя узами брака. Лас-Вегас, 7 февраля 2009 г.
Электрорезонаторная гитара Корпус из хромированного металла, одинарный диффузор Ø 240 мм и высотой 30 мм, крышка резонатора золотого цвета, гриф: красное дерево, гриф: венге, вставки на грифе: точки, масштаб: 628 мм, ширина порожка: 42 мм, 19 ладов, Клен / Эбеновое дерево … Металлический хромированный корпус. Одинарный конус Ø 240 мм высотой 30 мм. P0501 hyundai sonata
Но я могу без шума опустить действие на Charvel, так что мне это нравится. лучше. Это правда, что анкерный стержень не предназначен для регулировки высоты струны, но также верно и то, что вам нужно отрегулировать анкерный стержень (если он нужен), прежде чем настраивать высоту струны … Одноразовый табачный вейп
Мост Callaham Vintage Hardtail с усовершенствованными латунными седлами, предназначенный для вибрато в стиле Bigsby Flat Mount Это наш винтажный хардтейл, модифицированный для использования … Zfs создают разреженный звук
Странно, чудесно, красиво, снова странно. Гитары и другие гитары и все, что мне нравится.Аноним http://www.blogger.com/profile/07068253323945482476 noreply … Два блока масс m1 и m2 связаны пружиной жесткости k
1/2 ″ выше красного дерева.Я измерил эту высоту, вынув бриджевый звукосниматель. Там есть полка из красного дерева, доступная там, где ввинчивается звукосниматель, ее высота составляет 1 3/4 дюйма, следовательно, верх бланка из красного дерева. Штангенциркулем я мог измерить высоту клена там, и это, кажется, самая высокая точка. Quantstudio application biosystems
Это «Gretsch Tenor G9120» от Hawaii Music Supply на Vimeo, где собраны высококачественные видео и люди, которые их любят. Gretsch Tenor G9120. 8 лет назад.Dentemp walgreens
Действие ореха определяется высотой струны над первым ладом.Очевидно, гитары имеют разные размеры ладов, и все игроки носят лады в разных точках и с разной скоростью. 2000 dodge durango no bus no start
10 мая 2020 г. · Гитара Бо Диддли была сделана Gretsch специально для этого легендарного гитариста.Еще в 1959 году Бо Диддли подошел к Гретчу и попросил их сделать ему гитару с довольно необычными формами. То, что они придумали, как Billy Bo Jupiter Thunderbolt. Бо Диддли любил гитару и играл на ней долгое время. Стоматологические материалы Target
Gretsch G2220 Junior Jet Electric Bass Guitar II. Gretsch G2224 Junior Jet Electric Bass Guitar II — еще одна бас-гитара с короткой шкалой, которая чрезвычайно популярна в гитарном сообществе и не зря. Если вы какое-то время участвовали в гитарном сообществе, возможно, вы слышали о бренде Gretsch. Winnebago revel 4×4 2021
18 декабря 2020 г. · Регулировка действия на басу (который представляет собой высоту струн от грифа) является важной частью общей настройки инструмента.Это необходимо делать, когда инструмент новый. Кроме того, изменение температуры и влажности, а также изменение толщины струны могут повлиять на настройку баса и потребовать регулировки … Тактические приклады Mcmillan
Слушайте Gretsch на Spotify. The Pinstripes · Song · 2009. Мы и наши партнеры используем файлы cookie, чтобы персонализировать ваш опыт, показывать вам рекламу на основе ваших интересов, а также в целях измерения и анализа.Инструменты управления проектным циклом pdf
Magnuson supercharger jeep jl
Настольная пила Ryobi
Seriale turcesti biz
Limewash кирпич до и после
Toyota RAV4, неизменно являющийся самым продаваемым внедорожником, предлагает отделку для любого бюджета.Лидирующая по цене комплектация LE довольно проста, но high-end Limited может бросить вызов Lexus NX с его многочисленными роскошными функциями. Новая модель TRD Off-Road обещает улучшенные возможности при выезде на трассу, а ее прочный дизайн заставляет ее выглядеть как дома рядом с внедорожником Toyota 4Runner и пикапом Toyota Tacoma. Динамика вождения Ute имеет тенденцию к приземленному уровню, но большинство потребителей сочтут езду RAV4 2020 года комфортной, а его управляемость — приятной. Взрослые пассажиры будут довольны большим пространством для ног на передних и задних сиденьях.А грузовой отсек не уступает лучшим в своем сегменте. Если вам нужна экономия топлива, рассмотрим гибрид RAV4, который мы рассмотрим отдельно.
Что нового в 2020 году?Toyota представила модель TRD Off-Road на 2020 год, которая поставляется с 18-дюймовыми колесами для каждой модели, шинами повышенной проходимости, комплектом подвески TRD с красными винтовыми пружинами и множеством логотипов TRD, разбросанных по интерьеру. и экстерьер. Toyota добавила спутниковое радио SiriusXM и совместимость с Android Auto в стандартной комплектации.Модель XLE теперь оснащена сиденьем водителя с электроприводом, но люк в крыше уже не входит в стандартную комплектацию; вам нужно будет выбрать пакет «Удобство», если он вам нужен. Комплектация Adventure среднего класса получила обтянутые кожей рулевое колесо и ручку переключения передач, но задняя дверь с электроприводом теперь не является обязательной. А в комплектации Limited в стандартную комплектацию входит аудиосистема премиум-класса JBL.
Цены и какой купитьНам трудно устоять перед крутым новым TRD Off-Road, но мы бы проявили сдержанность и выбрали XLE Premium среднего класса.Он предоставляет множество стандартного оборудования, которого не предлагают LE и XLE, такого как 19-дюймовые колеса, задняя дверь с электроприводом, двухзонный автоматический климат-контроль, обивка из искусственной кожи, а также обтянутые кожей рулевое колесо и ручка переключения передач. Toyota делает свой набор функций помощи водителю стандартным для всей линейки, поэтому нет особых причин выбирать более дорогую комплектацию — если, конечно, вам действительно не нужны вентилируемые сиденья.
Двигатель, трансмиссия и характеристикиRAV4 2020 года использует 203-сильный двигатель 2.5-литровый четырехцилиндровый двигатель в паре с восьмиступенчатой автоматической коробкой передач. Передний привод остается стандартным, но теперь Toyota предлагает выбор систем полного привода в зависимости от отделки салона. Модель Adventure, на которой мы ездили, была оснащена системой полного привода, которая может отключать задний мост, когда это необходимо для повышения экономии топлива. На нашем тестовом треке RAV4 оказался быстрее своего предшественника: мы зафиксировали время разгона до 60 миль в час за 8,0 секунды с моделью 2019 года и 8,3 секунды с моделью 2018 года. Послушный четырехцилиндровый двигатель может быть шумным, особенно при проезде по шоссе. попытки, и трансмиссия на нашем тестовом автомобиле, казалось, спотыкалась, когда требовалось переключить на пониженную передачу.Гибрид RAV4, рассматриваемый отдельно, в наших руках показал еще более быстрое время разгона до 60 миль в час за 7,4 секунды. RAV4 чувствует себя прочным и стабильным при движении по межгосударственным трассам и сохраняет свои позиции на извилистом участке дороги, но он не внушает такой же уверенности, как наш фаворит в сегменте, Mazda CX-5.
Тойота
Экономия топлива и расход топлива в реальных условияхRAV4 получил конкурентоспособные в своем классе оценки экономии топлива от Агентства по охране окружающей среды.RAV4 с передним приводом могут проезжать по шоссе примерно до 35 миль на галлон. Полноприводное Adventure доставило нам 32 мили на галлон на нашем 200-мильном шоссе с экономией топлива, что на 1 милю на галлон меньше оценочного рейтинга EPA для этой отделки. Это ставит его в соответствие с Honda CR-V и представляет собой огромное улучшение по сравнению с моделью Adventure 2018 года (которая проработала всего 25 миль на галлон), но его превосходит Volkswagen Tiguan, который выдал 33 мили на галлон.
Интерьер, комфорт и грузВнутри RAV4 отдает предпочтение эргономике и компактному пространству для хранения вещей.Полезная полка охватывает приборную панель и дополняет большую центральную консоль. Версия, которую мы тестировали, отличалась сочетанием приятных на ощупь поверхностей и приятного пластика. Базовая отделка, названная LE, довольно спартанская и идет с тканевыми сиденьями, пластиковым рулевым колесом и однозонным ручным климат-контролем. На следующем шаге XLE Toyota добавляет несколько льгот, но в XLE Premium есть гораздо больше, что может понравиться, включая обивку из искусственной кожи, обтянутые кожей рулевое колесо и ручку переключения передач, а также прошитую и мягкую приборную панель и подлокотник центральной консоли.Задние сиденья RAV4 легко складываются, они складываются почти до плоского положения, открывая кабину для груза. Мы помещаем 10 наших ручных чемоданов за заднее сиденье и 22 — в сложенном виде. Nissan Rogue здесь соответствовал RAV4, но Subaru Forester превзошел его на единицу по каждому показателю.
Тойота
Информационно-развлекательная система и возможности подключенияНижняя отделка салона поставляется с 7,0-дюймовым сенсорным экраном, но 8,0-дюймовый ноутбук можно приобрести, начиная с уровня XLE.Информационно-развлекательная система включает в себя мобильную точку доступа 4G LTE и возможности Apple CarPlay и Android Auto. Аудиосистема премиум-класса JBL, навигация и беспроводная зарядная панель для смартфона не являются обязательными. Если вы выберете уровень отделки салона без навигации, не волнуйтесь. Все другие модели поставляются с функцией Scout GPS Link, которая позволяет совместимым смартфонам отображать карту и пошаговые инструкции на экране приборной панели.
Функции безопасности и помощи водителюНациональная администрация безопасности дорожного движения (NHTSA) присвоила RAV4 пятизвездочную оценку, а Страховой институт дорожной безопасности (IIHS) назвал его лучшим выбором безопасности.Линейка RAV4 может похвастаться множеством стандартных систем помощи водителю. Покупатели также могут добавить мониторинг слепых зон и предупреждение о перекрестном движении сзади, но эти функции требуют дополнительных затрат на базовой модели. Основные функции безопасности включают:
- Стандартное автоматическое экстренное торможение с обнаружением пешеходов
- Стандартное предупреждение о выезде с полосы движения с системой помощи при удержании полосы
- Стандартный адаптивный круиз-контроль
Гарантии Toyota соответствуют отраслевым линии с трехлетним покрытием от бампера до бампера и пятью годами для трансмиссии.Все модели поставляются с бесплатным плановым обслуживанием в течение двух лет — большая редкость в этом сегменте.
- Ограниченная гарантия распространяется на три года или 36000 миль
- Гарантия на трансмиссию распространяется на пять лет или 60000 миль
- Бесплатное обслуживание распространяется на два года или 25000 миль
Дополнительные функции и характеристики